Power, Sex, Suicide: Mitochondria and the Meaning of Life (30 page)
Read Power, Sex, Suicide: Mitochondria and the Meaning of Life Online
Authors: Nick Lane
Tags: #Science, #General
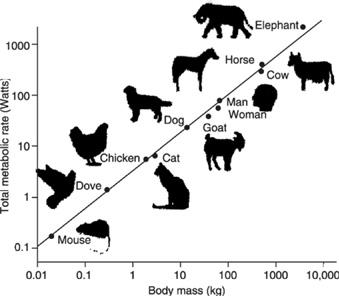
In considering dogs, of course, Rubner was considering only one species, even though the different breeds vary dramatically in size and appearance. Half a century later, the Swiss-American physiologist Max Kleiber plotted log metabolic rate against log mass of different species, and constructed his famous curve from mice to elephants. To his and everybody else’s surprise, the exponent was not 2/3 as expected, but 3/4 (0.75; or in actual fact 0.73, rounded up;
Figure 12
). In other words, for every three steps in the log metabolic rate, there were four steps in log mass. Other researchers, notably the American Samuel Brody, came to a similar conclusion. Even more unexpectedly, the 0.75 exponent turned out to apply to more than just the mammals: birds, reptiles, fish, insects, trees, even single-celled organisms, have all been placed on the same curve: metabolic rate is claimed to vary with the 3/4 power of mass (or mass
3/4
) across an extraordinary 21 orders of magnitude. Many other traits also vary with an exponent based on multiples of one quarter (such as 1/4 or 3/4), giving rise to the general term ‘quarter-power scaling’. For example, the pulse rate, the diameter of the aorta (even the diameter of tree trunks), and lifespan, all conform roughly to ‘quarter-power scaling’. A minority of researchers, most persuasively Alfred Heusner at UC Davis, have contested the universal validity of quarter-power scaling, but it has nonetheless entered virtually all standard biological texts as ‘Kleiber’s law’. It is often said to be one of the few universal laws in biology.
1
12
Graph showing the scaling of resting metabolic rate with body mass across mammals of widely differing mass, from mouse to elephant. The slope of the line on a log–log plot is ¾, or 0.75, which is to say that the line rises 3 steps up the vertical axis in the space of 4 steps on the horizontal axis. This slope gives the exponent. The metabolic rate is said to vary with the three-quarter power of mass, or mass
3/4
.
Why on earth metabolic rate should vary with the 3/4 power of mass remained a mystery for another half century; and indeed the glimmerings of an answer are only now beginning to materialize, as we shall see. But one point was patent: while a 2/3 exponent, connecting the metabolic rate with the surface area to volume ratio, made sense for warm-blooded mammals and birds, there was no obvious reason why it should apply to cold-blooded animals, such as reptiles and insects: they don’t generate heat internally (or at least not much), so the balance of heat generation and heat loss could hardly be the dominating factor. From this perspective a 3/4 exponent made as much, and as little, sense as a 2/3 exponent. But while various attempts have been made to rationalize the 3/4 exponent, none ever really convinced the whole field.
Then, in 1997, a high-energy particle physicist at Los Alamos National
Laboratory, Geoffrey West, joined forces with the ecologists James Brown and Brian Enquist, at the University of New Mexico, Albuquerque (through the Santa Fe Institute, an organization that fosters cross-disciplinary collaborations). They came up with a radical explanation based on the fractal geometry of branching supply networks, such as the circulatory system of mammals, the respiratory tubes of insects (the trachea), and the plant vascular system. Their densely mathematical model was published in
Science
in 1997, and the ramifications (if not the maths) swiftly captured the imagination of many.
Fractals (from the Latin
fractus
, broken) are geometric shapes that look similar at any scale. If a fractal is broken into its constituent parts, each part still looks more or less the same, because, as the pioneer of fractal geometry Benoit Mandelbrot put it, ‘the shapes are made of parts similar to the whole in some way’. Fractals can be formed randomly by natural forces such as wind, rain, ice, erosion, and gravity, to generate natural fractals, like mountains, clouds, rivers, and coastlines. Indeed, Mandelbrot described fractals as ‘the geometry of nature’, and in his landmark paper, published in
Science
, in 1967, he applied this approach to the question advanced in its title:
How Long is the Coast of Britain?
Fractals can also be generated mathematically, often by using a reiterative geometric formula to specify the angle and density of branches (the ‘fractal dimension’).
Both types of fractal share a property known as scale invariance, which is to say they ‘look’ similar whatever the magnification. For example, the contours of a rock often resemble those of a cliff or even a mountain, and for this reason geologists like to leave a hammer lying around in photographs, to enable viewers to grasp the scale. Similarly, the pattern of river tributaries looks alike for a vast continental system, such as the Amazon basin when viewed from space, or small streams seen from the top of a hill, or even soil erosion in the back garden from the bathroom window. For mathematical ‘iterative’ fractals, a repeating geometric rule is used to generate an infinite number of similar shapes. Even the most complex and beautiful fractal images, seen adorning T-shirts and posters, are built from reiterations of geometric rules (often quite complicated ones), followed by plotting the points spatially. For many of us, this is as close as we’ll ever get to the beauty of deep mathematics.
Most of nature’s fractals are not really true fractals, in that their scale invariance does not extend to infinity. Even so, the pattern of twigs on a branch resembles the branching of the tree as a whole; and the branching pattern of blood vessels in a tissue or an organ resembles that of the body as a whole—it can be difficult to grasp the scale. Stepping up again in scale, the cardiovascular
system of an elephant resembles that of a mouse, but the system as a whole is magnified by nearly six orders of magnitude (in other words, the cardiovascular system of an elephant is nearly a million times bigger than that of a mouse; a 1 followed by six zeros). When networks retain a similar appearance over such a scale, the natural language to describe them is fractal geometry; if nature’s branching networks are not true fractals, they are still close enough to be modelled accurately using these mathematical principles.
West, Brown, and Enquist asked themselves whether the fractal geometry of nature’s supply networks might account for the apparently universal scaling of metabolic rate with body size. This makes perfectly good sense, for the metabolic rate corresponds to the consumption of food and oxygen, and these don’t arrive at the individual cells of an animal by diffusing across the body surface, but by way of the branching supply network—blood vessels in our own case. If the metabolic rate is constrained by the delivery of these nutrients, it’s reasonable to assume that it should depend ultimately on the properties of the supply network. In their 1997
Science
paper, West, Brown, and Enquist made three basic assumptions. First, they assumed that the network serves the entire organism—it must supply all cells, and so fill the entire volume of the organism. Second, they assumed that the smallest branch of the network, the capillary, is a size-invariant unit, which is to say that all capillaries are the same size in all animals, regardless of the size of the animal. And third, they supposed that the energy necessary to distribute resources through the network is minimized: over evolutionary time, natural selection has optimized the supply network to deliver nutrients with the minimum time and effort.
A number of other factors, relating to the elastic properties of the tubes themselves, also needed to be considered, but we needn’t worry about these here. The upshot is this. To maintain a self-similar fractal network (one that ‘looks’ the same on any scale), while scaling up the body size over orders of magnitude, the total number of branches rises more slowly than the volume. This is observed to be true. For example, a whale is 10
7
(10 million) times heavier than a mouse, but has only 70 per cent more branches going from the aorta to the capillaries. According to the idealized calculations of fractal geometry, the supply network ought to take up relatively less space in a large animal, and so each capillary ought to serve a larger number of ‘end-user’ cells. Of course this means that cells must partition less food and oxygen between them; and if they receive less food to burn, then presumably they would be forced to have a slower metabolic rate. How much slower, exactly? The fractal model predicts that the metabolic rate should correspond to body mass to the power of ¾. Picture this as the slope of the line on a log-log plot: for every three steps in log metabolic rate, there are four steps in log mass. In other words, the fractal model calculates, from first principles, that the metabolic rate should scale with
mass
3/4
, thus explaining the universality of Kleiber’s law, the quarter-power scaling rule. If this is true, then the entire living world is subject to the rules of fractal geometry. They determine body size, population density, lifespan, speed of evolution—everything.
As if this weren’t enough, the fractal model goes still further and makes a radical general prediction. Because Kleiber’s law apparently applies not only to large organisms that manifestly have a branching supply network, like mammals, insects, and trees, but also to simpler creatures that seem to lack a supply network, like single cells, then they too must have some sort of fractal supply network. This is radical as it implies that there exists a whole echelon of biological organization that we have not yet detected, and even the proponents feel obliged to talk about a ‘virtual’ network, whatever that might be. Even so, many biologists are receptive to the possibility, for the cytoplasm is now seen to be far more organized than the amorphous jelly that is passed off in text books. The nature of this organization is elusive, but it is clear that the cytoplasm ‘streams’ through the cell, and that many biochemical reactions are more carefully defined in space than had been assumed. Most cells have a complex internal architecture, including branching networks of cytoskeleton filaments and mitochondria; but is this really a fractal network, that obeys the same laws of fractal geometry? While it unquestionably branches, there is a very modest resemblance to the
tree-like
network of circulatory systems (
Figure 13
). If fractal geometry applies to self-similar systems, these do not look similar.
To address such intangibles, West, Brown, and Enquist recast their model to eliminate the need for explicit structures, like a branching anatomy, grounding it instead in the geometry of hierarchical networks (networks embedded within networks, like a set of Russian dolls). Other physicists, notably Jayanth Banavar at the University of Pennsylvania and his colleagues, have attempted to simplify the network model to eliminate the need for fractal geometry altogether; but they, too, specify a branching supply network. Various abstruse mathematical arguments have filled the pages of prestigious science journals every few months since the late 1990s, often delivering withering mathematical ripostes, such as ‘this must be incorrect because it violates dimensional homogeneity…’ The debate tends to polarize biologists, who are only too aware of exceptions to supposedly universal rules (‘yeah, but what about the crayfish?’), and physicists, like West, who seek a single unifying explanation. West doesn’t mince his words: ‘If Galileo had been a biologist he would have written volumes cataloguing how objects of different shapes fall from the leaning tower of Pisa at slightly different velocities. He would not have seen through the distracting details to the underlying truth: if you ignore air resistance, all objects fall at the same rate regardless of their weight.’
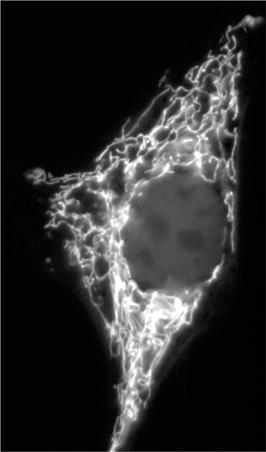
13
The mitochondrial network in a mammalian cell in a tissue culture, stained with the dye MitoTracker. Mitochondria often move around the cell, and can form into reticulated networks, as here, but these networks don’t really resemble a fractal tree.
West and Brown teamed up with the Los Alamos biochemist William Woodruff to report perhaps the most thought-provoking finding of all in 2002. They published data in the
Proceedings of the National Academy of Sciences
in which they extended their fractal model to the mitochondria. They showed that mitochondria, and even the thousands of miniscule respiratory complexes within individual mitochondria, could be plotted onto the same universal quarter-power scale. In other words, they said, the relationship between metabolic rate and body size extends from the level of individual respiratory complexes right up to
the blue whale, spanning ‘an astounding 27 orders of magnitude’. When I set out the proposal for this book, I had it in mind to discuss their paper. I had read it carefully enough to find the central argument compelling, but had not really come to grips with its implications. I’ve been struggling with them ever since—is it true that a straight line connects the metabolic rate of individual complexes within the mitochondria with the metabolic rate of a blue whale? And if it is true, what does it mean?