Eight Little Piggies (12 page)
Read Eight Little Piggies Online
Authors: Stephen Jay Gould
Unfortunately, we cannot experiment on extinct animals and have no direct evidence for quadrate hearing in the actual ancestors of mammals. However, we do know that reptilian quadrates can transmit sound while still acting as part of a jaw joint, for several modern reptiles use an important quadrate path to their inner ear. (These creatures are not mammalian ancestors to be sure, but they do demonstrate the possibility, indeed the actuality, of this crucial multiple modality in the evolution of mammalian hearing.) Snakes, for example, have no external ear or eardrum, and many scientists had considered them entirely deaf, until recent studies illustrated sensitivity to sound over most of the body, especially around the large lung that can transmit vibrations to the inner ear. But another route offers special advantages to a creature so close to the substrate by God’s direct decree: “…upon thy belly shalt thou go, and dust shalt thou eat all the days of thy life.” Snakes hear primarily by placing their heads on the ground and passing vibrations from the lower jaw to the quadrate and finally to the stapes—thus closely following the eventual mammalian pathway. In addition, direct experiments on several lizards and on the tuatara of New Zealand show that vibrations directed at the quadrate are passed to the stapes and recorded in the brain.
May I confess an ulterior motive in closing—for complex and abstract excursions can be mere glosses upon simpler aims. In-jokes have delicious qualities because they are inaccessible to all but the initiated. But sometimes, in-jokes are so good that we long to share them, yet despair for the volume of background required. Well, this essay can be read as nothing but an extended pony for understanding one of my favorite humorous poems. My colleague John Burns, a lepidopterist now in Washington but formerly at Harvard, used to introduce our weekly seminars with his punny doggerel. We loved the poems and came more to hear his introductions than to suffer through the subsequent speeches. John finally published his verses in a volume called
Biograffiti
(Demeter Press, 1975), with an introduction by yours truly. My favorite is a pithy epitome of mammalian ear evolution, entirely incomprehensible to 99 percent of the population, but now vouchsafed to you, my dear readers, as a small reward for your persistence and as a dessert after this ponderous dissertation:
Evolution of Auditory Ossicles
With malleus
Aforethought
Mammals
Got an earful
of their ancestors’
Jaw.
FIORELLO LA GUARDIA
may be destined to go down in history primarily as godfather to an airport. But he was a great mayor for New York in tough years of depression and war. (My birth certificate even bears his signature—well, at least a stamped version.) He also possessed in abundance the trait that we find most welcome, but encounter all too rarely, in people of accomplishment—a willingness to acknowledge occasional and inevitable error. In his most famous quip, La Guardia once remarked, “When I make a mistake it’s a beaut!”
Scientific “misconduct” is now a hot topic, both for journalists and members of Congress. In this somewhat frantic climate, we should pause to consider the essential distinction between fraud and error—for the two concepts are diametrically opposed, although self-appointed watchdogs sometimes make the tragic mistake of uniting them as graded forms of malfeasance. Fraud is a social and psychological pathology, although science must learn to police itself. Error is the inevitable byproduct of daring—or of any concentrated effort for that matter. You might as well legislate against urination after beer drinking.
No great work of science has ever been free of error, and any extensive or revolutionary work must contain a few of La Guardia’s beauts. Intellectual progress is a complex network of false starts and excursions into trial and error. Darwin’s
Origin of Species
, for example, sprinkles numerous errors into its ocean of reforming validity. The errors are so frequent, and so varied, that we might even try to establish categories.
Darwin, first of all, commits several errors of fact. Here I would skip the dull and quotidian misreporting of information and concentrate on the far more interesting errors based on predictions from theoretical premises that turn out to be false or exaggerated. Darwin’s commitment to gradualism, for example, led him to make at least two prominent, and outstandingly wrong, conjectures: (1) He gave a time of more than 300 million years for the “denudation of the Weald” (the erosion of the region, forty miles wide, between the north and south Chalk Downs in southern England), based on his belief in the steady, grain-by-grain character of geological erosion. But alteration need not proceed so slowly, or so continuously, and the actual time is one-third to one-fifth of Darwin’s generous allotment. (2) Multicellular animal life begins with geological abruptness at the “Cambrian explosion” some 550 million years ago. Darwin, who rejected biological rapidity even more zealously than the geological variety, predicted that the “explosion” must be illusory and that the pre-Cambrian history of multicellular animal life must be as long as, or longer than, the 570 million years of success ever since. We now have an excellent record of pre-Cambrian life—and no multicellular animals arise until just before the Cambrian explosion.
A second category might be labeled errors of judgment: political miscalculations really. The savvy Darwin made few mistakes in this mode, but he slipped occasionally by giving free rein to fatuous speculations in a treatise that gained its power by sinking a weighty anchor in sober fact and avoiding the fanciful conjectures of previous writing about evolution. In a passage that he would later rue, and that gave aid, comfort, rhetorical advantage, and belly laughter to the enemy, Darwin wrote:
In North America the black bear was seen by Hearne swimming for hours with widely open mouth, thus catching, like a whale, insects in the water…. If the supply of insects were constant, and if better adapted competitors did not already exist in the country, I can see no difficulty in a race of bears being rendered, by natural selection, more and more aquatic in their structure and habits, with larger and larger mouths, till a creature was produced as monstrous as a whale.
(Later editions of the
Origin
kept the first factual sentence and expunged all the rest.) A statement like this need not be false (indeed, as a speculation, we cannot tell); the important thing, as Machiavelli would have said, is to avoid the appearance of silliness.
A third category, perhaps the most revealing, includes mistakes that most of us don’t recognize because we make them ourselves. Call them errors of thoughtless convention. I include here those passive repetitions of standard cultural assumptions stated so automatically, or so deeply (and silently) embedded within the structure of an argument, that we scarcely detect their presence. Darwin may have been the greatest intellectual revolutionary of the nineteenth century, but he made a few outstanding errors in this category, most related to his ambiguity on the great bugbear of progress—a concept that had no place in the basic mechanics of natural selection, but that Darwin, as an eminent Victorian, could not abandon entirely.
Consider Darwin’s treatment of the evolution of vertebrate lungs and their relationship with the swim bladders of bony fishes—an example that Darwin obviously viewed as important to his general argument because he repeats the story half a dozen times in the
Origin
. Darwin begins by noting, correctly, that the lung and swim bladder are homologous organs—different versions of the same basic structure, just as a bat’s wing and a horse’s foreleg share a common origin indicated by the similar arrangement of bones in body parts that now work in such different ways. But Darwin then draws a false inference from the fact of homology. He claims, with increasing confidence ending in certainty, that lungs evolved from swim bladders:
All physiologists admit that the swim bladder is homologous…in position and structure with the lungs of the higher vertebrate animals; hence there seems to me to be no great difficulty in believing that natural selection has actually converted a swim bladder into a lung, or organ used exclusively for respiration. I can, indeed, hardly doubt that all vertebrate animals having true lungs have descended by ordinary generation from an ancient prototype, of which we know nothing, furnished with a floating apparatus or swim bladder.
Many readers will be puzzled at this point, as I have perplexed several generations of students by presenting the argument in this form. What can be wrong with Darwin’s claim? The two organs are homologous, right? Right. Terrestrial vertebrates evolved from fishes, right? Yes again. So lungs must have evolved from swim bladders, right? Wrong, dead wrong. Swim bladders evolved from lungs.
I love this example, especially as a pedagogical tool, because an outstandingly counterintuitive assertion—the evolution of swim bladders from lungs—becomes the favored hypothesis with sudden and stunning clarity as soon as we shed a common, disabling assumption and start considering the question in a different light. The problem lies with a chronic confusion—abetted by cultural prejudice in this case—between structural sequence and branching order.
The literature of experimental psychology often reports comparative data of performance on various tests for learning in, say, a planarian worm, a crab, a carp, a turtle, and a dog. These are often reported as an “evolutionary sequence” of mental advance. Such statements make evolutionary biologists howl in rage or, if our mood be better, merely with laughter. This motley crew of animals represents no evolutionary sequence at all: vertebrates did not arise from arthropods; mammals did not evolve from turtles; and carp are further from the fishes that did give rise to terrestrial descendants than aardvarks are from humans. However, although the psychologists are dead wrong in their terminology of “evolutionary order,” their sequence may have some validity as a structural series—worm, bug, fish, turtle, and dog might express some increasing property of neurological functioning.
When we turn to another common sequence—fish, amphibian, reptile, mammal, monkey, human—the problems intensify, for now we cannot even speak of a legitimate structural sequence. Frogs live in different places, but are they “higher” than sword-fishes or sea horses? What odds would you put on a ground sloth going up against a
Triceratops?
Fine, you say; no necessary progress here, but surely this venerable lineage records
the
path of vertebrate evolution. And now we come to the crux of the error about lungs and swim bladders. If this sequence is
the
path of vertebrate change, then swim bladders must evolve to lungs, as Darwin said—for the canonical fish, the first member of the series, has a swim bladder, while all of us at the top have lungs.
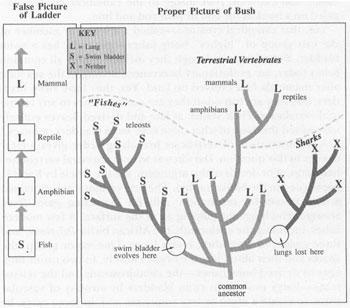
The correct sequence of lung evolving to swim bladder is almost inconceivable on the false evolutionary model of a ladder in vertebrate evolution. But, with the proper iconography of a bush, the sequence becomes clear.
Joe LeMonnier. Courtesy of
Natural History.
But we wallow in a double confusion when we make this “intuitively obvious” assertion—first, the false assumption of progress, which makes the lung a “higher” organ than a swim bladder and thus unfit for creatures on the bottom; second, and more seriously, the confusion of ladders and bushes, or sequences and branching orders. Fish-amphibian-reptile-mammal is not
the
road of change among vertebrates; it represents only one pathway among thousands in the complexly branching bush of vertebrate evolution (the accompanying figure should make my argument clear). All the other pathways lead to creatures that we continue to call “fish” in the vernacular. In terms of variety in anatomical design, we find far more diversity among the creatures called fish than among all the terrestrial vertebrates put together. The terrestrial line is a single branch, with astounding success to be sure, but with limited diversity in underlying anatomical structure (whatever the outward variety of flying birds, slithering snakes, and thinking people). By contrast, fishes are astoundingly disparate in basic design and include lineages that separated a hundred million years before any terrestrial vertebrate arose. Consider the jawless lampreys, the boneless sharks (also lacking either lung or swim bladder), and the odd coelacanth; don’t confine your image to the canonical creature impaled on a hook at the end of your rod and line.
Yes, that canonical creature—called a teleost, or member of the vast group of “higher” bony fishes—generally has a swim bladder. But teleosts, although they include almost all common fishes today, are evolutionary latecomers, arising in the sea long after mammals first evolved on land. Yes, they have swim bladders, and they are fish—but they are not ancestors to any terrestrial vertebrate. Their status, as late and derived, leaves entirely unresolved the issue of what came first: swim bladders or lungs.
A reconstruction of vertebrate branching order gives a clear answer to this question: Darwin was wrong; ancestral vertebrates had lungs. (For details of this argument, see the article by Karel F. Liem, cited in the bibliography). The first vertebrates maintained a dual system for respiration: gills for extracting gases from seawater and lungs for gulping air at the surface. A few modern fishes, including the coelacanth, the African bichir
Polypterus
, and three genera of lungfishes, retain lungs. One major group, the sharks and their allies, lost the organ entirely. In two major lineages of derived bony fishes—the chondrosteans and the teleosteans—lungs evolved to swim bladders by atrophy of vascular tissue to create a more or less empty sac and, in some cases, by loss of the connecting tube to the esophagus (called the trachea in humans and other creatures with lungs). Some fishes retain the connection of swim bladder with esophagus; they can inflate their swim bladders by gulping air at the surface. Fishes with separate swim bladders usually extract gases from blood flowing through an extremely fine and rich system of vessels surrounding the bladder and possessing one of the loveliest technical names in all biology—the
rete mirabile
, or “wondrous network.”
I would not wish to issue overt praise for mistakes, but Darwin’s error on the swim bladder falls into the category that we welcome as particularly instructive, for correction involves a sudden shift from the “can’t be” to the completely obvious—that almost thrilling property of scales falling from eyes. The agent of correction, moreover, is not a new and pristine fact, but a change in an underlying conceptual structure.
Let us then praise Darwin’s fruitful error on this basis, but also for another, and even more important, reason. Darwin may have gotten his sequence backward, but he was using the story to illustrate a vital and widely misunderstood principle of evolutionary theory—and the illustration works just as well whether swim bladders evolve to lungs or vice versa. Why, then, was Darwin so interested in this issue in the first place?
One common argument against evolution held (and still holds among the lingering opposition) that small changes within a “basic kind” might occur, producing the range from Chihuahua to Great Dane, or Shetland pony to old dobbin hauling the Budweiser truck. But transitions between types are forbidden because fundamental novelties cannot arise by evolution. The classic form of this argument holds that since “novel” structures often arise (or so evolutionists claim) from ancestral organs with strikingly different functions, transitional forms would be inviable because they would exist in the never-never land of utter unworkability, with one key function degenerated and another not quite established. To cite a classic case (with an elegant resolution as we shall see), how could reptiles evolve into mammals if bones that articulate the reptilian jaw must evolve to the malleus and incus (hammer and anvil) of the mammalian middle ear (see Essay 6)? No intermediary form could live without a jaw articulation, as the leisurely earward transition occurred. In other words, both the “before” and the “after” make sense as functional organisms, but the “in between” doesn’t work.