Full House (24 page)
McShea was then able to apply the third test of skewing to three vertebral dimensions measured in three lineages. The mean skew value for all nine distributions is actually negative (—.19), not significantly so to be sure, but quite a comeuppance for the traditional view of complexity as driven—a conclusion that implies positive (right) skewing for component lineages!
McShea then summarizes his entire study (1994, page 1761):
The minimum complexity of vertebral columns probably did not change (indeed, the actual minimum seems to have remained close to the theoretical minimum), ancestor-descendant comparisons in subclades of mammals revealed no branching bias, and the mean subclade skew was negative, all pointing to a passive system.
One study doesn’t prove a generality any more than a single swallow makes a summer, but when our first rigorous data point to a conclusion so at variance with traditional views, we must sit up and take notice, and then go out to make more tests. The few other available studies also support the passive rather than the driven mode. In an interesting report, presented at paleontological sessions at the 1995 annual meeting of the Geological Society of America in New Orleans, McShea provided the first results for a quite different meaning of complexity—developmental rather than morphological, and defined as the number of independent growth factors that build a structure through embryology (practically measured as correlation coefficients between pairs of measures, with perfect positive correlation indicating that the two measures represent only one mode of growth, and zero correlation implying that the two measures indicate different developmental influences).
Working with Benedikt Hallgrimsson and Philip D. Gingerich, McShea applied this method to a large series of classical and excellent data on measurements of fossil teeth, compiled over many years by Gingerich on evolutionary sequences for several mammalian lineages in the Bighorn Basin of Wyoming. They found no trend to increasing complexity and concluded: "Tes
t
s detected no bias, no tendency for non-hierarchical developmental complexity either to increase or decrease."
In the only other comprehensive study, using an interesting metric for complexity applied to a very different group of organisms, Boyajian and Lutz (1992 and personal communication) studied one of the classic examples of supposedly driven evolution toward greater complexity—and again found evidence only for the passive mode!
Ammonites are extinct relatives of the modern chambered nautilus— coiled cephalopod shells housing animals related to modern squid and octopuses. Internal chambers meet the external shell at a boundary known as a "suture line." In nautiloids the suture line is usually straight or mildly wavy, but in ammonites the suture line can become intricately sinuous and digitated. In the everyday sense that sinuous and digitated looks more complicated than straight or mildly wavy, an old paleontological truism asserts that ammonite sutures become more complex through time. Ever since the earliest days of paleontology, increasing complexity of the ammonite suture has ranked among the two or three "classic" trends that "everybody knows" in the fossil record of invertebrates.
Boyajian and Lutz used a clever measure of "fractal dimension" to assess the complexity of ammonite sutures. (Heretofore, the trend has merely been asserted subjectively, rather than proven quantitatively, because no one could figure out a rigorous way to measure the complexity of such a twisty line.) Fractals have become a hot topic of popular culture, but in a technical sense, fractals are curves and surfaces that exist between ordinary dimensions. Since a straight line has a fractal dimension of one, and a plane a fractal dimension of two, twisty lines must measure between one and two—that is, between a minimum of one for the straight line between two points, and the unattainable maximum of two for a line that twists and turns so much that it fills an entire plane between the two points at opposite edges. The higher the fractal dimension, the more "complex" the suture in our visceral and traditional sense that the squiggliest lines are most elaborate. Boyajian and Lutz measured the fractal dimension of a suture in each of 615 genera of ammonites spanning the full range of their history. The measured scope of fractal dimensions runs from just a tad over 1.0 (very simple sutures are close to straight lines) to just over 1.6 for the most complex.
All early ammonites grew fairly simple sutures, and some measure near the theoretical minimum of 1.0 for a straight line. Thus, as in McShea’s vertebrae, where founders showed minimal complexity, any movement away from initial values could only be upward! This origin at a true left wall set the supposedly driven trend to increasing sutural complexity— for many later ammonites have very complex sutures, and scientific imagination can always drum up some putative adaptive reason for why complex sutures should be better, and therefore favored by natural selection (greater shell strength against hydrostatic pressure, and increasing area for attachment of muscles, have been favored).
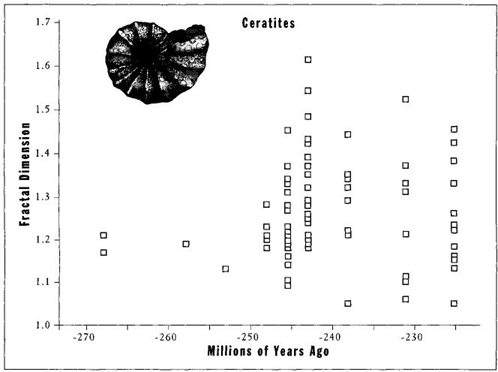
FIGURE 35 Evolution of complexity as measured by fractal dimension in a group of ammonites (the ceratites). The first species (left) have simple sutures near the left wall. Low values persist and even decrease through the group’s history, but variation also expands into the only open direction of higher fractal dimension.
But Boyajian and Lutz could find no evidence for a driven trend; all data identify the trend as probably passive—an incidental effect of minimally simple beginnings at a left wall, followed by no bias whatever for increasing complexity in individual lineages thereafter. Most lineages of ammonoids maintain species of low complexity throughout their history (see Figure 35 for an example). Most important, Boyajian and Lutz found no bias for increasing complexity among all the ancestor-descendant pairs they could specify (note the similarity to Arnold et al.’s discovery of no bias for increasing size in ancestor-descendant pairs of forams—see page 161). Finally, if complexity is such a good thing, then genera with more complex sutures should live longer. But Boyajian and Lutz found no correlation between sutural complexity and geological longevity (see Figure 36).
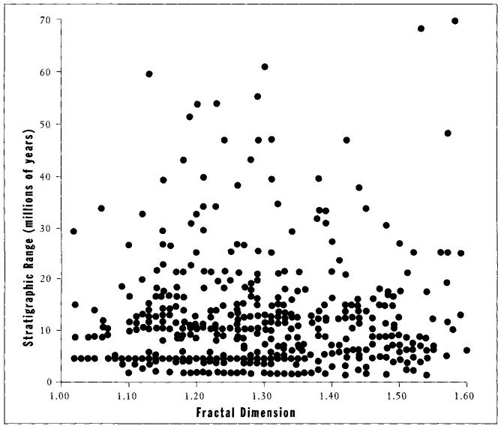
FIGURE 36 Longevity (in millions of years) for ammonite genera on the vertical axis plotted against fractal dimension on the horizontal axis. There is no correlation between complexity and success as measured by longevity.
Only the most minuscule proportion of scientific studies ever gets reported in the press, and these decisions often bear little correlation with the importance of such studies for professionals. Better relationships can be found between the decision to report and the degree to which a conclusion disturbs conventional notions (often misconceptions) about the nature of things. The research of McShea and Boyajian is important to professionals, but their studies also received rare and extensive coverage in the popular press because they challenged something that "everybody knows"—and that turns out to be probably wrong: the supposed drive to increasing complexity as the defining feature of life’s evolutionary thrust. Consider the leads to the two major press accounts. Carol K. Yoon in The New York Times (March 30, 1993):
Surveying life’s rich parade from the first single-celled beings in the primordial soup to the diverse array of organisms into which they developed, evolutionary biologists have marveled at the ever more complex cast of creatures that has continued to grace the planet. The evolution of larger brains, more efficient metabolisms and more elaborate social systems all seem to support the conventional wisdom that complexity increases during evolution. So clear is the trend that some biologists suggest that the evolutionary process is actually driving the increase in complexity.... But in two of the first studies to measure these trends, based on mammals’ backbones and fossil shells, researchers say they have been unable to detect any overall evolutionary drive toward greater complexity.
And Lori Oliwenstein in Discover (June 1993):
Everybody knows that organisms get better as they evolve. They get more advanced, more modern, and less primitive. And everybody knows, according to Dan McShea (who has written a paper called "Complexity and Evolution: What Everybody Knows"), that organisms get more complex as they evolve. From the first cell that coalesced in the primordial soup to the magnificent intricacies of Homo sapiens, the evolution of life—as everybody knows—has been one long drive toward greater complexity. The only trouble with what everybody knows ... is that there is no evidence it’s true.
Few intellectual tyrannies can be more recalcitrant than the truths that everybody knows and nearly no one can defend with any decent data (for who needs proof of anything so obvious). And few intellectual activities can be more salutary than attempts to find out whether these rocks of ages might crumble at the slightest tap of an informational hammer. I love the wry motto of the Paleontological Society (meant both literally and figuratively, for hammers are the main tool of our trade): Frango ut patefaciam—I break in order to reveal.
A Note on the Fatal Weakness of the Last Straw
People under assault, and hopelessly overmatched, often do the opposite of what propriety might suggest: they dig in when they ought to accommodate. We call this behavior "siege mentality." Davy Crockett, Jim Bowie, and Co. won posthumous immortality by their intransigence at the Alamo, but an honorable surrender (given their hopeless situation and the certainty of carnage with continued fighting) might have secured the more worldly privilege of telling good war stories over a beer at a Texan bar (for independence from Mexico would have been won in any case) some twenty years later.
I believe that the power of arguments against inherent progress as a driving force of evolution, and the strength of data on the modal bacter and the passive character of trends to the right tail, must now evoke something like a siege mentality among those who still wish to maintain that evolution validates the primacy and domination of human life on our planet. To what may such people now turn as a source of natural solace? The modal bacter must be acknowledged as dominant by any reasonable criterion. The right tail exists, but only as a little appendage that cannot wag the dog of life’s full house. Moreover, the right tail arose as an incidental consequence, in a passive trend powered by constraints of life’s origin next to the left wall—and not by any cause or bias that favored increasing complexity as a natural good, and a driving thrust of evolution.
The embattled traditionalist must therefore stand his ground on the right tail of his natural habitat. He must adopt a siege mentality and dig in to protect his own restricted turf. The right tail, he must now admit, may be small and merely consequential. But grant me, he pleads, this last potential natural comfort: "May I not at least be a king in my own restricted castle? I once thought that my domain extended over all nature—that all else must be viewed as predictably preparatory to my eventual origin. I am now prepared to admit the hubris and falsity of this view. I reside on a small and incidental tail. But I am, at least, the creature of maximal complexity (’properly’ defined by neural elaboration) on this tail, which I therefore dominate by right. This right tail, however passively fashioned, still had to develop and, ultimately, had to spawn a creature like me. Give me, then, at least, this one remaining solace in a parody of a fine old song: ’It had to be me, wonderful me; it had to be me.’
"Let me, in short, live like Pio Nono (the nineteenth-century Pope Pius IX). My predecessors held temporal power over much of Europe. I once ruled a good part of Italy, though I am now confined to a tiny principality—Vatican City—within Rome. But at least my rule here is absolute— and I can proclaim my infallibility!"
But even this reverie—a bit manic to be sure, for embattlement tends to inspire paranoia and delusions of limited grandeur—cannot be sustained. The claim that a conscious creature like us must evolve because we can predict the development of an expanding right tail for all of life represents a classic "category mistake"—in this case, the false inference of a particular from a valid generality. The right tail did predictably arise (if only as a passive consequence), but any individual creature on the right tail of earthly life at this particular time represents a fortuitous and improbable result, one actualization among a hundred million unrealized alternatives. Wind back the tape of life to the origin of modern multicellular animals in the Cambrian explosion, let the tape play again from this identical starting point, and the replay will populate the earth (and generate a right tail of life) with a radically different set of creatures. The chance that this alternative set will contain anything remotely like a human being must be effectively nil, while the probability of any kind of creature endowed with self-consciousness must also be extremely small.
This theme of radical contingency and improbability for particulars, whatever the predictability of general patterns (with humans clearly defined as an improbable particular, not part of any expected generality), does not fall under the scope of this book. But I do need to epitomize the argument at this point (as abstracted from my previous book Wonderful Life) because the traditional view, challenged and upended by contingency, forms the last refuge for a hope that we might validate human supremacy as an expected result of general evolutionary principles.
Under the traditional model of evolutionary history as a "cone of increasing diversity," life moves ever upward to greater progress, and outward to a larger number of species—from simple Cambrian beginnings for multicellular animals to our modern levels of progress and range of diversity. Under this iconography, pathways actually followed run along predictable courses that would be at least roughly repeated in any replay. But a radically different view, suggested by a thorough restudy of soft-bodied fossils in the Burgess Shale and other Cambrian faunas, indicates that an inverted iconography may be more appropriate—with maximal anatomical range of disparity reached early in life’s history, followed by the extinction of most initial experiments and the "settling down" of life’s diversity to just a few of the original possibilities. Moreover, we have strong reason to suspect that the loss of most, and survival of just a few, occurred more through a distribution of lottery tickets than by victories for predictable cause based on higher levels of progress among winners. In the "pure" lottery model, "tickets" are distributed at random and few initial lineages receive such a blessing. Any replay distributes the tickets to another random set, and leads to a radically different group of survivors. Since our own lineage of vertebrates held a tenuous position among these initial experiments—with only two early Cambrian precursors known as fossils, Pikaia from the Burgess Shale, and Yunnanozoon recently described from Chengjiang in China (see Chen et al., 1995, and Gould, 1995)—we must assume that most replays would not include the survival and flourishing of vertebrates. All of us—from sharks to rhinos to humans—would then have been excluded from the history of life,
If this good fortune of radical contingency occurred but once, with predictability based on progress prevailing thereafter, then we might view human emergence as close to inevitable following one lucky spin of fortune’s wheel. But radical contingency is a fractal principle, prevailing at all scales with great force. At any of a hundred thousand steps in the particular sequence that actually led to modern humans, a tiny and perfectly plausible variation would have produced a different outcome, making history cascade down another pathway that could never have led to Homo sapiens, or to any self-conscious creature.
If one small and odd lineage of fishes had not evolved fins capable of bearing weight on land (though evolved for different reasons in lakes and seas), terrestrial vertebrates would never have arisen. If a large extraterrestrial object—the ultimate random bolt from the blue—had not triggered the extinction of dinosaurs 65 million years ago, mammals would still be small creatures, confined to the nooks and crannies of a dinosaur’s world, and incapable of evolving the larger size that brains big enough for self-consciousness require. If a small and tenuous population of protohumans had not survived a hundred slings and arrows of outrageous fortune (and potential extinction) on the savannas of Africa, then Homo sapiens would never have emerged to spread throughout the globe. We are glorious accidents of an unpredictable process with no drive to complexity, not the expected results of evolutionary principles that yearn to produce a creature capable of understanding the mode of its own necessary construction.