I Have Landed (44 page)
Authors: Stephen Jay Gould
By contrast, the preformationists rallied behind a story of differentiation that envisaged all the structural complexity of the neonate as already present within the initial cell, and only brought to visibility during embryology. In the caricatured version, preformationism has usually been ridiculed as the belief that a perfect homunculus lies within each sperm or egg cell. No serious scientific preformationist held such a view. Rather, they argued that all structures must be present in the initial cell, but in too tiny, too transparent, and too diffuse a state to be visible (like the chaos at the outset of Genesis 1, and not like a fully formed homunculus). Embryology then becomes a differentiative process of concentration, coagulation, solidification, and growth.
When evolutionary ideas pervaded embryology in the nineteenth century, the two leading interpretations continued to uphold contrasting stories of addition or differentiation. Haeckel's famous theory of recapitulation held, in a purely additive account, that sequential steps in embryonic complexification repeated the evolutionary accretion of successive adult stages to the ancestral lineageâso that a complex animal, in its embryology, literally climbed its own family tree. The primary alternative, von Baer's theory of differentiation, argued that the visual simplicity of an early stage does not represent an ancient ancestor that must then be augmented (as in Haeckel's additive theory), but
rather a more general form of greater homogeneity and lesser differentiation, holding all potential for the definitive complexity that eventually develops in each lifetime. Thus, at an early stage of development, we know that the embryo will become a vertebrate, then (at a later stage) a mammal, then a primate, then a hominoid, and finally a human beingâa process of increasingly finer specification, contrasted with Haeckel's additive model of ever-increasing complexity in accretion.
When we consider the other historical process that led to our human formâthe much longer evolutionary construction
of Homo sapiens
in geological time, rather than the embryological generation of each individual
Homo sapiens
in nine monthsâwe note that concepts of evolution may also be classified into additive and differentiative models. Darwinism embodies an additive view. Because the Darwinian style of explanation has prevailed within science, we tend to forget that several abandoned theories of evolution advocated differentiative models. These accounts imagined that the first vertebrate in the Cambrian explosionâa boneless and jawless creature just an inch or two in lengthâalready contained all the parts and potentials that evolution would necessarily elaborate into human form in a distant future. The supposed mechanisms for such a “programmed” differentiation spanned the full gamut from God's direct actions (in a few overtly theological accounts) to principles embodied in unknown, but entirely physical, laws of nature (for some atheistic versions at an opposite speculative extreme).
If allegiance to an additive or differentiative model implied no consequences for skewing our views of life in disparate directions, then we could dismiss the entire subject as an effete intellectual game without meaning for scientific understanding. But our preferred theories often act as biases that strongly influence our basic conceptions of the natural worldâand additive versus differentiative views of historical sequences do not hold the same intellectual weights, properties, and implications. We might summarize the differences, looking at lessons from the history of science, by saying that each basic model features a defining property and struggles with a major problem.
Stories of differentiation work primarily from the inside out. That is, the sequence begins with all eventual results already preformed, albeit unexpressed, within an initial homogeneity. How, then, can this potential become actualized? In explicit contrast, stories of addition operate primarily from the outside in. That is, the sequence begins with truly unformed stuff, promiscuous potential that could be drawn into any number of pathways by outside forces. How, then, can such a gloppy mass be carved by external agents into such an exquisitely complex final product (an even worse problem for embryology
than for evolution, because the carving must follow the same basic path each time for normal embryos within a species, whereas each evolutionary result arises only once).
In the major weight of difference between the two models, stories of differentiation fit better for determined systems in a predictable world, whereas stories about addition hold the conceptual edge in a contingent world where each historical sequence may follow innumerable (and unpredictable) options, with the actual result conditioned by the particular set of external prods that a rolling ball of promiscuous potential happens to encounter in its trajectory through time. For this primary reason, our modern embryological models tend to be primarily differentiative, and our evolutionary models primarily additive.
After all, embryology does generally follow an internally prescribed route specified not by the preformed parts of preformationists, but by the programmed instructions of modern genetic understanding. (We should not accuse the eighteenth-century preformationists of stupidity for placing the right idea into the wrong substance. After all, their intellectual world did not include a concept of programmed information, except, perhaps, as embodied in the old trifle of music boxes or the newfangled invention of the Jacquard weaving loomâwhereas no sentient person in our age of genes and computers could fail to assimilate such informational models as an intellectual centerpiece.)
By contrast, the evolution of any lineage wanders along contingent and unpredictable paths of a uniquely complex history. The few lineages, including our own, that do become more complex through time may add their increments of sophistication in a sequence that makes sense after the fact. But even an omniscient observer could never designate, for certain, the next step in an unpredictable future. Therefore, as a description of evolution, additive models that introduce sequential steps from the outside work better than differentiative models that must hypothesize an entire future as already implicit and enfolded within any current form.
Under this analysis, we should not be surprised that Genesis 1, despite our usual and unconsidered readings, tells a tale of differentiation rather than addition. After all, if God proceeded with the usual care and thought conventionally attributed to his might, he probably had a pretty accurate idea about the finished product even before he began the work. Biological evolution, on the other hand, at least as viewed under the limits of our eminently fallible mental machinery, seems to wander along a wondrously erratic set of specific pathways within its broad predictabilities.
Our preferred intellectual models do make a difference, and we must therefore be sensitive to the disparate implications of additive and differentiative
models as we struggle to understand the history of life. Still, I think that any passionate and sentient person can feel the same emotional thrill that emanates from either intellectual interpretation. We live in one helluva fascinating universe, whatever its modalities of construction. Thus, if I may beg one last indulgence from my readersâthis time for ending with the same image that I invoked in essay 19 for a different treatment of Genesis and evolutionâI happily embrace the common sentiment behind two maximally different views of organic order: the differentiative model of Genesis 1, with its ending of sublime satisfaction: “And God saw every thing that he had made, and, behold, it was very good.” And the additive model of natural selection, so lovingly described by Charles Darwin in the last paragraph of
The Origin of Species:
“There is grandeur in this view of life.”
Â
Â
ARSING
AND
P
ROCEEDING
21
Linnaeus's Luck?
C
AROLUS
L
INNAEUS
(1707â1778),
THE
FOUNDER
OF
MOD
ern taxonomy and the focus of this essay, frequently cited an ancient motto to epitomize his view of life:
natura non facit saltum
(nature does not make leaps). Such unbroken continuity may reign in the material world, but our human passion for order and clear distinction leads us to designate certain moments or events as “official” beginnings for something discrete and new. Thus the signatures on a document define the birth of a nation on July 4, 1776, and the easily remembered eleventh hour of the eleventh day of the eleventh month (November 11, 1918) marks the armistice to a horrible war supposedly fought to end all contemplation of future wars. In a small irony of history, our apostle of natural continuity also became the author and guardian of a symbolic leap to noveltyâfor the modern taxonomy of animals officially began with the publication of the definitive tenth edition of Linnaeus's
Systema Naturae
in 1758.
The current classification of animals may boast such a formally recognized inauguration, but an agreement about beginnings does not guarantee a consensus about importance. In fact, the worth assigned to taxonomy by great scientists has spanned the full range of conceivable evaluations. When Lord Rutherford, the great British physicist (born in New Zealand), discovered that the dates of radioactive decay could establish the true age of the earth (billions rather than millions of years), he scorned the opposition of paleontologists by branding their taxonomic labors in classifying fossils as the lowest form of purely descriptive activity, a style of research barely meriting the name “science.” Taxonomy, he fumed, could claim no more intellectual depth than “stamp collecting”âan old canard that makes me bristle from two sides of my being: as a present paleontologist and a former philatelist!
Rutherford's anathema dates to the first decade of the twentieth century. Interestingly, when Luis Alvarez, a physicist of similar distinction, became equally enraged by some paleontologists during the last decade of the twentieth
century, he invoked the same image in denigration: “they're not very good scientists; they're just stamp collectors.” I continue to reject both the metaphor and the damning of all for the stodginess of a majorityâfor Luis had exploded in righteous and legitimate frustration at the strong biases that initially led most paleontologists to reject, without fair consideration, his apparently correct conclusion that the impact of a large extraterrestrial body triggered the mass extinction of dinosaurs and about 50 percent of marine animal species 65 million years ago.
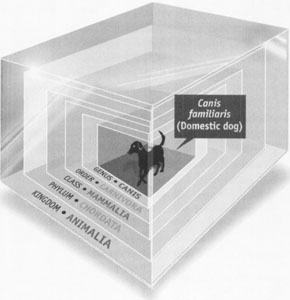
Linnaean taxonomy in one of its geometric portrayals as boxes within boxes for continually finer specifications
.
The phony assumption underlying this debasement of taxonomy to philately holds that the order among organisms stands forth as a simple fact plainly accessible to any half-decent observer. The task of taxonomy may then be equated with the dullest form of catalogingâthe allocation of an admittedly large array of objects to their preassigned places: pasting stamps into the designated spaces of nature's album, putting hats on the right hooks of the world's objective hatrack, or shoving bundles into the proper pigeonholes in evolution's storehouse, to cite a standard set of dismissive metaphors.
In maximal contrast, the great Swiss zoologist Louis Agassiz exalted taxonomy as the highest possible calling of all, when he opened Harvard's Museum of Comparative Zoology in his adopted land in 1859. Each species, Agassiz argued, represents the material incarnation on earth of a single and discrete idea in the mind of God. The natural order among speciesâtheir taxonomyâtherefore reflects the structure of divine thought. If we can accurately identify the system of interrelationships among species, Agassiz concluded, we will stand as close as rationality can bring us to the nature of God.
Notwithstanding their maximally disparate judgments of taxonomy, Rutherford and Agassiz rank as strange bedfellows in their shared premise that a single objective order exists “out there” in the “real world,” and that a proper classification will allocate each organism to its designated spot in the one true system. (For Rutherford, this order represents a basically boring and easily ascertainable aspect of macroscopic natureâtoo far removed from the atomic world of fundamental laws and causes to generate much scientific interest or insight. For Agassiz, in greatest conceivable contrast, this order represents our best shot for grasping the otherwise arcane and inaccessible intellect of God himself.)
In framing a modern “Goldilocks” defense for the importance of taxonomyâfar warmer than Rutherford's icy indifference, but not quite so hot as Agassiz's impassioned embraceâwe must begin by refuting their shared assumption that one true order exists “out there,” and that correct classifications may be equated with accurate maps. We can best defend the scientific vitality
of taxonomy by asserting the opposite premise, that all systems of classification must express theories about the causes of order, and must therefore feature a complex mixture of concepts and perceptsâthat is, preferences in human thinking combined with observations of nature's often cryptic realities. Good taxonomies may be analogized with useful maps, but they reveal (as do all good maps) both our preferred mental schemes and the pieces of external reality that we have chosen to order and depict in our cartographic effort.