In Search of Memory: The Emergence of a New Science of Mind (13 page)
Read In Search of Memory: The Emergence of a New Science of Mind Online
Authors: Eric R. Kandel
Tags: #Psychology, #Cognitive Psychology & Cognition, #Cognitive Psychology
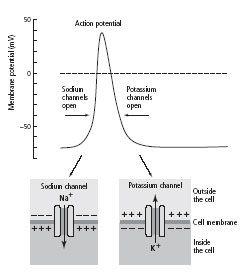
5–5 The Hodgkin-Huxley model of the intracellularly recorded action potential.
The influx of positively charged sodium ions (Na+) changes the cell’s internal voltage and produces the upstroke of the action potential. Almost immediately, potassium channels open and potassium ions (K+) flow out of the cell, producing the downstroke and returning the cell to its original voltage.
When the neuron is at rest, the voltage-gated channels are shut. When a stimulus reduces the cell’s resting membrane potential sufficiently, say from-70 millivolts to-55 millivolts, the voltage-gated sodium channels open and sodium ions rush into the cell, creating a brief but dramatic surge in positive charge that moves the membrane potential from-70 millivolts to +40 millivolts. In response to the same change in membrane potential, the sodium channels close after a split second’s delay and the voltage-gated potassium channels open briefly, increasing the outflow of positive potassium ions and rapidly returning the cell to its resting state of-70 millivolts (figure 5–5).
Each action potential ultimately leaves the cell with more sodium on the inside and more potassium on the outside than is optimal. Hodgkin found that this imbalance is remedied by a protein that transports excess sodium ions out of the cell and potassium ions back into the cell. Eventually, the original concentration gradients of sodium and potassium are reestablished.
Once an action potential has been generated in one region of the axon, the current it generates excites the neighboring region to generate an action potential. The resulting chain reaction ensures that the action potential is propagated along the entire length of the axon, from the site where it was initiated to its terminals near another neuron (or muscle cell). In this way a signal for a visual experience, a movement, a thought, or a memory is sent from one end of the neuron to the other.
For their work, now known as the ionic hypothesis, Hodgkin and Huxley shared the Nobel Prize in Physiology or Medicine in 1963. Hodgkin later said the prize should have gone to the squid, whose giant axon made their experiments possible. But this modesty overlooks the extraordinary insights that the two men provided—insights that gave the scientific community, including new converts like me, confidence that we could understand signaling in the brain at a deeper level.
WHEN MOLECULAR BIOLOGY WAS APPLIED TO BRAIN SCIENCE
, it revealed that the voltage-gated sodium and potassium channels are actually proteins. These proteins span the width of the cell membrane and contain a fluid-filled pathway, the ion pore, through which ions pass. Ion channels are present in every cell of the body, not just neurons, and they all use essentially the same mechanism Bernstein proposed to generate the resting membrane potential.
Much as the neuron doctrine had done earlier, the ionic hypothesis strengthened the link between the cell biology of the brain and other areas of cell biology. It offered the final proof that nerve cells can be understood in terms of the physical principles common to all cells. Most important, the ionic hypothesis set the stage for exploring the mechanisms of neuronal signaling on the molecular level. The generality and predictive power of the ionic hypothesis unified the cellular study of the nervous system: it did for the cell biology of neurons what the structure of DNA did for the rest of biology.
In 2003, fifty-one years after the ionic hypothesis was formulated, Roderick MacKinnon at Rockefeller University received the Nobel Prize in Chemistry for obtaining the first three-dimensional picture of the atoms that form the protein of two ion channels—a non-gated potassium channel and a voltage-gated potassium channel. Several features revealed by MacKinnon’s highly original structural analysis of these two proteins had been predicted with amazing prescience by Hodgkin and Huxley.
SINCE THE MOVEMENT OF IONS THROUGH CHANNELS IN THE
cell membrane is critical to the functioning of neurons, and the functioning of neurons is critical to mental functioning, it is not surprising that mutations in the genes that code for ion channel proteins should lead to disease. In 1990 it became possible to pinpoint the molecular defects responsible for human genetic diseases with relative ease. Shortly thereafter, several ion channel defects that underlie neurological disorders of muscle and of the brain were identified in rapid succession.
These disorders are now referred to as channelopathies, or disorders of ion channel function. For example, a disorder called familial idiopathic epilepsy, an inherited epilepsy of newborns, has been found to be associated with mutations in genes coding for a potassium channel. Recent progress in exploring channelopathies and the development of specific treatments for them can be attributed directly to the large body of basic scientific knowledge about ion channel function already on hand, thanks to Hodgkin and Huxley.
I
arrived in Harry Grundfest’s laboratory in 1955 in the wake of a major controversy about how neurons communicate with one another. Hodgkin and Huxley’s epochal work had solved the long-standing mystery of how electrical signals are generated within neurons, but how does signaling occur
between
neurons? For one neuron to “talk” to the next neuron in line, it would have to send a signal across the synapse, across the gap between the cells. What kind of signal could that be?
Until they were proven wrong in the early 1950s, Grundfest and other leading neurophysiologists firmly believed that this small signal across the gap between two cells was
electrical
, the result of the flow into the postsynaptic neuron of the electrical current produced by an action potential in the presynaptic neuron. But starting in the late 1920s, evidence began to accumulate that the signal between certain nerve cells might be
chemical
in nature. This evidence came from studies of nerve cells in the autonomic, or involuntary, nervous system. The autonomic nervous system is considered part of the peripheral nervous system because the bodies of its nerve cells lie in clusters, called peripheral autonomic ganglia, which are located just outside the spinal cord and brain stem. The autonomic nervous system controls vital involuntary actions such as heart rate, blood pressure, digestion, and the modulation of breathing by emotion.
The new evidence gave rise to the chemical theory of synaptic transmission and led to a controversy humorously referred to as “soup versus spark,” with the “sparkers,” such as Grundfest, believing that synaptic communication is electrical and the “soupers” maintaining that it is chemical.
THE CHEMICAL THEORY OF SYNAPTIC TRANSMISSION AROSE
from studies by Henry Dale and Otto Loewi. In the 1920s and early 1930s, they investigated the signals sent by the autonomic nervous system to the heart and certain glands. Working independently, they discovered that when an action potential in a neuron of the autonomic nervous system reaches the terminals of the axon, it causes a chemical to be released into the synaptic cleft. That chemical, which we now call a neurotransmitter, moves across the synaptic cleft to the target cell, where it is recognized and captured by specialized receptors on the outer surface of the target cell membrane.
Loewi, a German-born physiologist living in Austria, examined the two nerves, or bundles of axons, that control heart rate: the vagus nerve, which slows it down, and the accelerans nerve, which speeds it up. In a crucial experiment on a frog, he stimulated the vagus nerve, causing it to fire action potentials that led to a slowing of the frog’s heart rate. He quickly collected the fluid around the frog’s heart during and just after stimulating the vagus nerve and injected that fluid into the heart of a second frog. Remarkably, the second frog’s heart rate also slowed down! No action potential had been fired to slow the second frog’s heart rate; instead, some substance that was released by the first frog’s vagus nerve transmitted the heart-slowing signal.
Loewi and the British pharmacologist Dale went on to show that the substance released by the vagus nerve is the simple chemical acetylcholine. Acetylcholine acts as a neurotransmitter, slowing the heart by binding to a specialized receptor. The substance released by the accelerans nerve to speed up heart rate is related to adrenaline, another simple chemical. For providing the first evidence that the signals sent across synapses from one neuron to another in the autonomic nervous system are carried by specific chemical transmitters, Loewi and Dale shared the Nobel Prize in Physiology or Medicine in 1936.
Two years after winning the Nobel Prize, Loewi personally experienced the disdain that the Austrian Nazis had for science and scholarship. The day after Hitler drove into Austria to the cheers of millions of my fellow citizens, Loewi was thrown in jail because he was a Jew. A scientist who had been professor of pharmacology at the University of Graz for twenty-nine years, Loewi was released two months later on the condition that he transfer his share of the Nobel Prize, still in a bank in Sweden, to a Nazi-controlled bank in Austria and leave the country immediately. He did so, moving to the New York University Medical School, where years later I had the privilege of hearing him lecture about his discovery of chemical signaling in the heart.
Loewi and Dale’s pioneering work on the autonomic nervous system convinced many neural scientists with a pharmacological bent that the cells in the central nervous system probably also use neurotransmitters to communicate across the synaptic cleft. However, some electrophysiologists, John Eccles and Harry Grundfest among them, remained skeptical. They acknowledged the importance of chemical transmission in the autonomic nervous system, but they were convinced that signaling between cells in the brain and spinal cord was simply too quick to be chemical in nature. They, therefore continued to favor the theory of electrical transmission in the central nervous system. Eccles hypothesized that the current produced by an action potential in the presynaptic neuron crosses the synaptic cleft and enters the postsynaptic cell, where it is amplified, leading to the firing of action potentials.
AS METHODS OF RECORDING ELECTRICAL SIGNALS CONTINUED
to improve, a small electrical signal was discovered at the synapse between motor neurons and skeletal muscle, proving that the action potential in the presynaptic neuron does not directly initiate the action potential in the muscle cell. Rather, the presynaptic action potential prompts a much smaller, distinctive signal, known as the synaptic potential, in the muscle cell. Synaptic potentials proved to be different from action potentials in two ways: they are much slower and their amplitude can vary. Thus on a loudspeaker like Adrian’s, a synaptic potential would sound like a soft, slow, protracted hissing rather than the sharp bang! bang! bang! of an action potential, and its volume would vary. The discovery of the synaptic potential proved that nerve cells use two different kinds of electrical signals. They use the action potential for long-range signaling, to carry information from one region of the nerve cell to another, and they use the synaptic potential for local signaling, to convey information across the synapse.
Eccles immediately recognized that synaptic potentials are responsible for Sherrington’s “integrative action of the nervous system.” At any given moment, a cell in any neural pathway is bombarded by many synaptic signals, both excitatory and inhibitory, but it has only two options: to fire or not to fire an action potential. Indeed, the fundamental task of a nerve cell is integration: it sums up the excitatory and inhibitory synaptic potentials it receives from presynaptic neurons and generates an action potential only when the total of excitatory signals exceeds the total of inhibitory signals by a certain critical minimum. Eccles saw that it is the capability of nerve cells to integrate all the excitatory and inhibitory synaptic potentials from the nerve cells that converge on it that assures the singleness of action in behavior that Sherrington described.
By the mid-1940s both sides of the debate agreed that a synaptic potential occurs in all postsynaptic cells and that it is the critical link between the action potential in the presynaptic neuron and the action potential in the postsynaptic cell. Yet that discovery just focused the argument more sharply: Was the synaptic potential in the central nervous system initiated electrically or chemically?
Dale and his colleague William Feldberg, another émigré from Germany, provided a key breakthrough when they discovered that acetylcholine, used in the autonomic nervous system to slow the heart, is also released by motor neurons in the spinal cord to excite skeletal muscles. This finding piqued the interest of Bernard Katz in exploring whether acetylcholine was responsible for the synaptic potential in skeletal muscle.
A prize-winning medical student at the University of Leipzig, Katz had fled Hitler’s Germany in 1935 because he was Jewish. He went to England and joined A. V. Hill’s laboratory at University College, London. Katz arrived at the English port of Harwich that February without a passport, which was, he remembered, “a terrifying experience.” Three months after his arrival, Katz attended a meeting in Cambridge where he had a ringside seat at a soup-versus-spark dustup. “To my great astonishment,” he would later write, “I witnessed what seemed almost a stand-up fight between J. C. Eccles and H. H. Dale, with the chairman, [Lord] Adrian, acting as a most uncomfortable and reluctant referee.” John Eccles, the leader of the sparkers, had presented a paper vigorously disputing a central claim of Henry Dale, the leader of the soupers, and his colleagues: that acetylcholine acts as a transmitter of signals at synapses in the nervous system. “I had some difficulty in following the argument, as I was not fully acquainted with the terminology,” Katz recalled. “The word
transmitter
conveyed to me something to do with radio communications, and as this did not make sense, the matter was a bit confusing.”
Indeed, Katz’s confusion aside, one of the problems with chemical transmission was that no one knew how an electrical signal in the presynaptic terminal could cause the release of a chemical transmitter and how that chemical signal could then be converted into an electrical signal in the postsynaptic neuron. Over the next two decades Katz joined the effort to address these two questions and to extend the work of Dale and Loewi from the autonomic nervous system to the central nervous system.
However, as with Hodgkin and Huxley, the threat of war interrupted Katz’s work. In August 1939, one month before World War II broke out, Katz, feeling uncomfortable as a German alien in London, accepted an invitation from John Eccles to join him in Sydney, Australia.

6–1
Three pioneers of synaptic transmission worked together in Australia during World War II and then went on to make major contributions individually. Stephen Kuffler (left, 1918–1980) characterized the properties of the dendrites of crayfish, John Eccles (middle, 1903–1997) discovered synaptic inhibition in the spinal cord, and Bernard Katz (right, 1911–2002) uncovered the mechanisms of synaptic excitation and of chemical transmission. (Courtesy of Damien Kuffler.)
As it happened, Stephen Kuffler, another scientist who left Europe to escape the Nazis and who also greatly influenced my thinking, also ended up in Sydney and joined Eccles’s laboratory (figure 6–1). A Hungarian-born, Viennese-trained physician-turned-physiologist, Kuffler left Vienna in 1938 because, in addition to having one Jewish grandfather, he was a socialist. Kuffler, a junior tennis champion in Austria, later joked that the real reason Eccles had asked him to join the laboratory was that he needed a skilled tennis partner. Although Eccles and Katz were vastly more experienced scientists, Kuffler amazed them with his surgical skills. He could dissect out individual muscle fibers to study the synaptic input from one motor axon to one muscle fiber, a real tour de force.
KATZ, KUFFLER, AND ECCLES SPENT THE WAR YEARS TOGETHER
arguing about chemical versus electrical transmission between nerve cells and muscle. Eccles attempted to reconcile the evidence of chemical transmission, which he insisted must be a slow process, with the rapidity of nerve-muscle signaling. He hypothesized that the synaptic potential has two components: an initial, rapid component mediated by an electrical signal, and a prolonged, residual action mediated by a chemical transmitter such as acetylcholine. Katz and Kuffler became newly minted soupers when they discovered evidence that the chemical acetylcholine is responsible even for the initial component of the synaptic potential in muscle. In 194f, after World War II had ended, Katz returned to England and Kuffler immigrated to the United States. In 1945 Eccles accepted a major professorship and moved to the University of Dunedin, in New Zealand, to set up a new laboratory.
As experiments cast more and more doubt on the electrical theory of synaptic transmission, Eccles, a large, athletic, and normally energetic and enthusiastic man, became dispirited. After we became friends in the late 1960s, he recalled how, in this despondent state, he underwent a great intellectual transformation for which he was forever grateful. That transformation occurred in the faculty club of the university, where Eccles went regularly for a break after the day’s work. On one such occasion in 1946, he met Karl Popper, the Viennese philosopher of science who had immigrated to New Zealand in 1937 anticipating that Hitler might annex Austria. In the course of their conversation, Eccles told Popper about the chemical-electrical transmission controversy and how he appeared to be on the losing side of a long and, for him, fundamental argument.
Popper was fascinated. He assured Eccles that there was no reason for despair. On the contrary, he urged him to be jubilant. No one was challenging Eccles’s research findings—the challenge was to his theory, his interpretation of the research findings. Eccles was doing science at its best. It is only when the facts become clear and competing interpretations of them can be brought into sharp focus that opposing hypotheses can clash. And only when sharply focused ideas clash can one of them be found wrong. Being on the wrong side of an interpretation was unimportant, Popper argued. The greatest strength of the scientific method is its ability to disprove a hypothesis. Science proceeds by endless and ever refining cycles of conjecture and refutation. One scientist proposes a new idea about nature and then other scientists work to find observations that support or refute this idea.