In The Blink Of An Eye (14 page)
Read In The Blink Of An Eye Online
Authors: Andrew Parker
Taking our tools to the Cambrian
Now that my palaeontological tour has entered the twenty-first century, it is safe to return to the Cambrian. Chapter 1 referred to the exceptional preservation of the Cambrian Burgess Shale fossils. The preservation of fine details has led to precise classifications. Limb parts and trace fossils are preserved in the Burgess Shale, and now these can be extrapolated to bring the fossils to virtual life. In fact the Burgess Shale and other Cambrian fossil assemblages have paved the way for wonderful ecological models to be constructed for Cambrian communities as a whole. Greatly exaggerated in size, the newly constructed Burgess scene within the Royal Tyrrell Museum in Alberta, Canada, includes a walk through a Cambrian reef where animals interact all around us, above and below. Here, the palaeontological techniques discussed in this chapter are brought to a crescendo as the Cambrian comes to virtual life.
It was the conditions of their burial that destined the Burgess organisms to make scientific headlines, and to star in the detailed Tyrrell model. A combination of an ideal clay substrate with the right cations, pH, and carbon content possibly engulfed the living Burgess organisms to preserve at least some of them in the wonderful condition in which we find them today. Original organic material from the Burgess organisms has been preserved in at least some cases. Nick Butterfield of Cambridge University demonstrated this fact with his delicate separation of organic
parts from the Cambrian rock. Acid was used to dissolve the rock matrix, where the fossils remained unharmed and simply floated away in the solution. These separated parts will be examined later in this book.
parts from the Cambrian rock. Acid was used to dissolve the rock matrix, where the fossils remained unharmed and simply floated away in the solution. These separated parts will be examined later in this book.
The Tyrrell model affirms that, as details of the Cambrian fauna are revealed at finer levels, the business of reconstructing Cambrian scenes is getting increasingly serious. The pioneer watercolours depicting Cambrian reef scenes, that for decades graced the corridors of natural history museums around the world, are making way for the sophisticated work of palaeo-artists. The crowded aquarium-like scenes of the amateur âwindows into the past' are becoming museum pieces themselves (quite literally). New reconstructions portray detailed movement in three dimensions among natural, spacious environments, as scientific principles are adhered to. X-ray photographs are revealing muscle attachment sites on the skeletons of Cambrian animals. Just as muscles were added externally to the first-century Jewish skull in lifelike proportions, now they are being added internally to the limbs of Cambrian arthropods - animals with exoskeletons like shrimps. When a skeleton is given virtual muscles in correct proportions, it can move naturally on the computer screen. Trilobite antennae are becoming conceptualised as flexible structures that can fold under the torso, as the body plates glide over each other and the animal rolls into a ball when danger approaches. On extension of the body, the gill plates are now considered to hang down quickly from the vaulted exoskeleton and flap in a style optimal for breathing when it is safe to do so. If all members of a community are brought to virtual life in this way, interactions between individuals and even entire food webs begin to manifest themselves. And work on Cambrian fossils and their reconstructions is accelerating.
In the dozen or so years since Stephen Jay Gould wrote
Wonderful Life
, advances in Cambrian biology have been considerable. The once âbizarre appearances of problematic species' have now been more closely linked to living species, following the discovery of new, intermediate forms that fill the gaps. The once mysterious long, thin bodies, of
Hallucigenia
and
Microdictyon
, with spindly legs, have now been placed within the velvet worm phylum of animals. Velvet worms exist today with thicker, worm-like bodies and stumpy legs. New velvet
worms have been uncovered from the Burgess Shale which share some important characteristics, such as claws, with living species and with
Hallucigenia
and
Microdictyon
. So the evolutionary holes have been filled.
Wonderful Life
, advances in Cambrian biology have been considerable. The once âbizarre appearances of problematic species' have now been more closely linked to living species, following the discovery of new, intermediate forms that fill the gaps. The once mysterious long, thin bodies, of
Hallucigenia
and
Microdictyon
, with spindly legs, have now been placed within the velvet worm phylum of animals. Velvet worms exist today with thicker, worm-like bodies and stumpy legs. New velvet
worms have been uncovered from the Burgess Shale which share some important characteristics, such as claws, with living species and with
Hallucigenia
and
Microdictyon
. So the evolutionary holes have been filled.
A variety of trace fossils have been found in Early Cambrian rocks. These include branching and spiralling burrows, and U-shaped and more complex migration paths through the sediment. The surface of the sea floor has preserved the trails and resting traces of creatures that walked and glided over the sediment. These are the footprints of animals with elaborate body forms and behaviours, including animals that were the first to
walk
on this planet, with tiny yet huge and historically significant steps.
walk
on this planet, with tiny yet huge and historically significant steps.
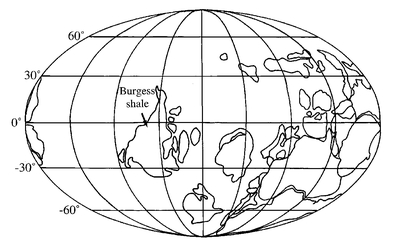
The biological indicators of environment within the Burgess Shale imply a tropical reef setting. But today the Burgess Shale is found halfway up a mountain in a snow-covered part of Canada - the furthest one can get from a tropical reef. Now it is understood that mountains today were perhaps once marine reefs, as a result of movements in the Earth's plates. In fact we can construct a world atlas so accurate that it could have been used to navigate in the Cambrian, at the time the Burgess organisms lived.
So the Burgess animals inhabited a near-equatorial position on the globe, hence their tropical environment. Now we would appear to know almost everything about their private lives, although surely the next decade will prove to be just as enlightening as the last. Remember the lesson learnt from the president of the Inventors' Association and his wisdom? But today we have amassed enough information, from ambiguous and esoteric signs of life flattened in shale, to consider the Burgess fossils as living organisms interacting within an ecosystem. And interactions between individuals from different species will prove to be important later in this book.
This chapter has demonstrated how our pictures of life on Earth through geological time have been constructed. By moving back through time gradually, and filling the gaps along the way, we can be less fearful of reconstructing ecosystems from an epoch as distant as the Cambrian. The logarithmic-style time travel employed in this chapter perhaps settles the nerves all the more - to begin with, it's nice to make
reconstructions of ancient but recent periods, periods we can test. Now we can be confident of the wealth of biological information we have extracted from Cambrian fossils, but this information ceases abruptly and simultaneously in all parts of the world as we split rocks formed before the Cambrian, beyond that auspicious borderline of 543 million years ago. It is no longer appropriate to extrapolate linearly and assume, with Darwin, that fossils of multicelled life with hard, external parts must exist in Precambrian rocks, implying that we simply haven't found them. Our fossil finds since Darwin's days have increased a hundredfold. But we are still without Precambrian signs of the characteristic external parts of animals today.
reconstructions of ancient but recent periods, periods we can test. Now we can be confident of the wealth of biological information we have extracted from Cambrian fossils, but this information ceases abruptly and simultaneously in all parts of the world as we split rocks formed before the Cambrian, beyond that auspicious borderline of 543 million years ago. It is no longer appropriate to extrapolate linearly and assume, with Darwin, that fossils of multicelled life with hard, external parts must exist in Precambrian rocks, implying that we simply haven't found them. Our fossil finds since Darwin's days have increased a hundredfold. But we are still without Precambrian signs of the characteristic external parts of animals today.
Figure 2.4
Palaeo-map of the world, at the time of the Burgess dynasty, showing the original location of the Burgess reef.
Palaeo-map of the world, at the time of the Burgess dynasty, showing the original location of the Burgess reef.
The characteristics of the fossil record through time have been assessed using quite convincing analytical methods, where the past 540 million years were examined. Although ancient rocks preserve less information on average than more recent rocks, the fossil record since the Cambrian explosion provides uniformly good documentation of the life of the past, and there is no reason not to extend this trend into the
Precambrian. So we are still without Precambrian fossils of the variety of body forms expressed in phyla today, other than the sponges, comb jellies and cnidarians. It seems certain that our modern view of animal evolution, and the Cambrian explosion, is correct. Equally, despite the numerous mass extinctions and recovery from those events
since
the Cambrian, new animal phyla have not evolved. These conclusions have been strengthened with every fossil discovery.
Precambrian. So we are still without Precambrian fossils of the variety of body forms expressed in phyla today, other than the sponges, comb jellies and cnidarians. It seems certain that our modern view of animal evolution, and the Cambrian explosion, is correct. Equally, despite the numerous mass extinctions and recovery from those events
since
the Cambrian, new animal phyla have not evolved. These conclusions have been strengthened with every fossil discovery.
In addition to presenting the evidence for Chapter 1, it is to be hoped that this chapter has supported a statement from John Maynard Smith's
The Theory of Evolution
: âThe study of fossils . . . can be made to reveal the way of life of animals now extinct.' We have ascertained how animals ran, swam, flew and burrowed in previous times. We have deduced their feeding habits, their daily activities and their favourite pastimes (almost). But after all this detail extracted from the fossil record, after all the constructions of virtual lifestyles, virtual climates and entire virtual ecosystems, there is still something missing from our interpretations of the past - colour. Is this a serious omission? It is time to examine colour in life today.
The Theory of Evolution
: âThe study of fossils . . . can be made to reveal the way of life of animals now extinct.' We have ascertained how animals ran, swam, flew and burrowed in previous times. We have deduced their feeding habits, their daily activities and their favourite pastimes (almost). But after all this detail extracted from the fossil record, after all the constructions of virtual lifestyles, virtual climates and entire virtual ecosystems, there is still something missing from our interpretations of the past - colour. Is this a serious omission? It is time to examine colour in life today.
3
The Infusion of Light
Whenever colour has been modified for some special purpose, this has been, as far as we can judge, either for direct or indirect protection, or as an attraction between sexes
CHARLES DARWIN,
On the Origin of Species
(first edition, 1859)
On the Origin of Species
(first edition, 1859)
Â
Â
Â
A series of Victorian doorways, staircases and corridors within Oxford University's Museum of Natural History eventually lead to the step of a more humble entrance in the far corner of the Gothic-style building. This is the door to the Huxley Room. Beyond this door lies an historic roof - its timbers absorbed the first words of evolution ever to be spoken to the public, during the Great Debate of 1860. Here, Thomas Huxley matched Bishop Wilberforce blow for blow in the original âscience versus religion' showdown. Huxley was defending Darwin's
On the Origin of Species
, published seven months earlier, in an attempt to prevent âsentiment interfering with intellect'. Darwin himself was absent, but Huxley skilfully succeeded in his task, and evolution began its infusion into the global language. It is worth pausing at the door to the Huxley Room.
On the Origin of Species
, published seven months earlier, in an attempt to prevent âsentiment interfering with intellect'. Darwin himself was absent, but Huxley skilfully succeeded in his task, and evolution began its infusion into the global language. It is worth pausing at the door to the Huxley Room.
After the Great Debate, the Huxley Room became moulded into an entomological collection room - it was filled with preserved insects. The last Victorian curator of the insect collection at Oxford's Natural History Museum, Sir Edward Bagnall Poulton, became fatefully attracted to the beetles within.
Poulton opened the door to the Huxley Room one morning, and as usual he took time to appreciate the architecture. Streams of sunlight,
illuminating the gently sloping sides of the roof and the many decorated beams, cut into the darkness of the room. He passed down the aisle of the Huxley Room, created by the two rows of wooden entomological cabinets. A drawer that had been removed from its cabinet for some time caused him to pause during his general inspection. The drawer was struck by the sun's rays, which were streaming through the round lens of a leaded window and became focused into a beam. Poulton blew away the dust from the glass lid and his eye, which had adapted to the intermittent darkness of the room, was at once arrested by a jewel. The beam of sunlight had ignited the metallic-blue colour of a carrion beetle, about the size of a thumbnail. The label, attached to the pin supporting the specimen, read â
Oiceoptoma
, Sumatra, Wallace 1866'. It was fitting that a specimen collected by Alfred Russel Wallace, a cofounder of the theory of evolution, should be found gleaming in a room where the theory had first been put on trial. Indeed, Darwin had collected other specimens here, but it was in the colour of that Wallace specimen that Poulton's real interest lay. Soon Poulton was placing all the entomological drawers under the beams of sunlight, which reflected rainbows on to the Huxley timbers, those pillars of evolutionary learning.
illuminating the gently sloping sides of the roof and the many decorated beams, cut into the darkness of the room. He passed down the aisle of the Huxley Room, created by the two rows of wooden entomological cabinets. A drawer that had been removed from its cabinet for some time caused him to pause during his general inspection. The drawer was struck by the sun's rays, which were streaming through the round lens of a leaded window and became focused into a beam. Poulton blew away the dust from the glass lid and his eye, which had adapted to the intermittent darkness of the room, was at once arrested by a jewel. The beam of sunlight had ignited the metallic-blue colour of a carrion beetle, about the size of a thumbnail. The label, attached to the pin supporting the specimen, read â
Oiceoptoma
, Sumatra, Wallace 1866'. It was fitting that a specimen collected by Alfred Russel Wallace, a cofounder of the theory of evolution, should be found gleaming in a room where the theory had first been put on trial. Indeed, Darwin had collected other specimens here, but it was in the colour of that Wallace specimen that Poulton's real interest lay. Soon Poulton was placing all the entomological drawers under the beams of sunlight, which reflected rainbows on to the Huxley timbers, those pillars of evolutionary learning.
Poulton eventually published a classification of colour in animals, and became âthe centre of gravity of entomological research in the British Empire'. He inspired a century of research on animal colouration and, in some ways, the clues that can be sought from this chapter towards solving the Cambrian enigma.
Before the VictoriansSome millennia earlier, Egyptians spoke of the âSun God'. They elevated the dung beetle to the status of higher being as it symbolically rolled sun-shaped objects around the desert. This âscarab' beetle was believed to represent the sun god Khepri, and in the Egyptian language the word âkheper' means both scarab beetle and existence. The Romans shared a similar interest in sunlight, though not only in a religious context. Heliography is the Roman art of signalling using the sun's rays
reflected on metal shields. It was sometimes employed to dazzle enemies, when sunlight was directed momentarily into their eyes. A flash of light is more conspicuous than a steady light, but at close range it can have a stunning effect. From an aeroplane, the reflection of sunlight from a car windscreen is extremely conspicuous, if not blinding. Unfortunately for the Romans, they were foiled by their own technology - Archimedes later engaged metal shields to concentrate sunlight on to the sails of invading Roman ships, causing them to burst into flames.
reflected on metal shields. It was sometimes employed to dazzle enemies, when sunlight was directed momentarily into their eyes. A flash of light is more conspicuous than a steady light, but at close range it can have a stunning effect. From an aeroplane, the reflection of sunlight from a car windscreen is extremely conspicuous, if not blinding. Unfortunately for the Romans, they were foiled by their own technology - Archimedes later engaged metal shields to concentrate sunlight on to the sails of invading Roman ships, causing them to burst into flames.
Exceptional intensities of light are also experienced in the natural world. If sunlight can be focused to cause materials to burn, imagine the effect it will have on a retina. And as the retina has been a product of evolution, so has retinal destruction. On this subject, angelfish become Hell's angelfish when territory is at stake.
Other books
The Coincidence 04 The Probability of Violet and Luke by Jessica Sorensen
My Secret Guide to Paris by Lisa Schroeder
An Illicit Temptation by Jeannie Lin
His Darkest Embrace by Juliana Stone
Boy Proof by Castellucci, Cecil
Horsing Around by Nancy Krulik
The Sea Taketh (Alex Singer) by Teresa Rae
Pyrus by Sean Watman
Sense of Wonder: A Century of Science Fiction by Grossman, Leigh
Your Scandalous Ways by Loretta Chase