The Flamingo’s Smile (21 page)
Read The Flamingo’s Smile Online
Authors: Stephen Jay Gould
TO MANY OUTSIDERS
, Indianapolis is nothing but one weekend a year and 500 miles of auto racing. In continuous reality, it is an attractive city filled with modern amenities and a liberal sprinkling of those older structures that unite our frenetic and uncertain present with a more comforting past. Last week, on a break from stated duties, I wandered through the Murat Temple of the Shrine and the enormous cathedral of Scottish Rite Masonry. These lodges must once have dominated the social life of Indianapolis; they may yet, for all I know, be important. But their gigantic buildings look forlorn and abandoned—cavernous Victorian rooms in dark wood and stained glass, dimly lit by available light, filled with old, overstuffed chairs occupied rarely by a few elderly men in odd-shaped hats. Surely, the old order changeth.
I was in Indianapolis to attend the annual meeting of the Geological Society of America.
*
There I watched, listened, and joined the debate as a group of my colleagues in paleontology began to dismantle an old order of thinking about old objects—and to construct a new and striking approach to a major feature of life’s history on earth: mass extinctions.
Paleontologists have known about mass extinctions from the inception of our science as a modern discipline. We have used them to mark the major divisions of our geological time scale—the boundaries between eras. The Permian extinction that rang out the Paleozoic era eliminated half the families of marine invertebrates; the Cretaceous extinction, marking the transition from Mesozoic to Cenozoic eras, wiped out some 15 percent of marine families, along with the most popular of all terrestrial creatures, the dinosaurs.
Nonetheless, though we have always acknowledged the reality of these great dyings, we have tried, in a curious way, to mitigate their effects, probably because our strong biases for gradual and continuous change force us to view mass extinctions as anomalous and threatening. We have, in short, attempted to depict mass extinction as a simple, quantitative extension of the slower disappearance, species by species, that characterizes normal times—larger and more abrupt, to be sure, but basically just more of the same. We have pursued two principal strategies to temper mass extinctions and bring them into harmony with events of ordinary times. First, we have emphasized continuity across the boundaries by trying to find direct ancestors for new forms that appear after an extinction among species that flourished just before the event. Second, we have toted the numerical patterns of extinctions to argue that the peaks were neither high nor abrupt enough to support a catastrophic view—in other words, we have argued that pulses of extinction were preceded by gradual declines lasting for millions of years, and that the peaks themselves do not stand so noticeably above the “background” rates of normal times.
Both these traditions were strongly challenged in Indianapolis in a series of separate and ostensibly unconnected papers that point to a common conclusion: mass extinctions must, by four criteria, be reinterpreted as ruptures, not the high points of continua. They are more
frequent
, more
rapid
, more
profound
(in numbers eliminated), and more
different
(in effect versus the patterns of normal times) than we had ever suspected. Any adequate theory of life’s history will have to treat them as special controlling events in their own right. They will not be fully explained by the evolutionary theory we have constructed for interaction among organisms and populations of normal times—that is, by nearly all conventional evolutionary theory now available.
Adolf Seilacher, professor of geology at Tübingen in Germany, presented the centerpiece of this unplanned assault upon tradition. Dolf is the greatest observer I have ever had the privilege of knowing. He looks at common objects, scrutinized by generations of researchers, and invariably sees something new and unexpected. This time he turned his superior gaze upon the oldest of all metazoan (multicellular animal) assemblages—the Ediacaran fauna. His paper offered a fundamental reinterpretation of these fossils, complete with wide-ranging implications for the entire history of life—and I sat spellbound as wave after wave of expanded meaning cascaded over me.
About 570 million years ago, our modern fossil record began with the greatest of geological bangs—the Cambrian explosion. Within a few million years, nearly all major groups of invertebrates with hard parts made their first appearance in the fossil record. For fully three billion years before, life had included little more than a long sequence of bacteria and blue-green algae. But the fossil record of early life does include one important, if last-minute, exception—first discovered in Australia but now known throughout the world—the Ediacaran fauna (named for the major Australian locality). In rocks just predating the Cambrian explosion, we find a moderately diverse assemblage of medium to large (up to a meter in length), soft-bodied, shallow-water marine invertebrates.
In the continuationist tradition that I identified above as a first strategy for softening the impact of mass extinctions, paleontologists have constantly tried to identify the Ediacaran animals with modern groups. Thus, the Ediacaran animals have been interpreted as jellyfish, corals, and worms—a continuity of evolutionary relationship across the greatest of all geological boundaries. Yet, as I argue in the following essay, the traditional ploy of forcing old and problematical fossils into modern taxonomic categories often fails badly. We must recognize that the early history of life should be studded with failed experiments—small groups that never achieved much diversity and bear only distant relationship with any modern animal. We might expect that our oldest fauna should contain a large number of such curiosities—yet all Ediacaran animals have been shoehorned, often with considerable effort, into modern groups.
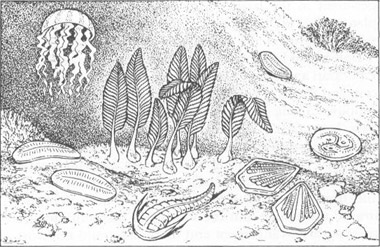
A conventional reconstruction of Ediacaran animals—my own, I regret to say. They are depicted as the ancestors of modern forms—jellyfish, soft corals, and worms.
FROM
A View of Life
BY S.E. LURIA, S.J. GOULD, AND S. SINGER. AFTER AN ILLUSTRATION IN
The History of Life
(1977)
BY A.E. LEE MCALESTER; REPRINTED BY PERMISSION OF PRENTICE-HALL, INC., ENGLEWOOD CLIFFS, N.J.
Dolf Seilacher now argues, turning the old view completely on its head, that the Ediacaran fauna contains no ancestors for modern organisms, and that every Ediacaran animal shares a basic mode of organization quite distinct from the architecture of living groups. The entire Ediacaran fauna, in other words, represents a unique and extinct experiment in the basic construction of living things. Our planet’s first fauna was replaced after a mass extinction, not simply improved and expanded.
Dolf began by showing that the traditional similarities of Ediacaran and modern animals are misleading and superficial, and that the Ediacaran forms could not work as their supposed living counterparts. Nearly all Ediacaran fossils have been falsely fit into three modern groups: jellyfish, corals, and segmented worms. Living jellyfish move by contracting a prominent ring of concentric muscles located at the outer edge of their bell; radial grooves for gathering and transporting food lie within the concentric muscles, toward the center. But the so-called Ediacaran medusoids reverse this arrangement and therefore could not work in the same way: concentric structures surround the center, and radial grooves lie on the outside.
Modern alcyonarian corals (“soft” corals, or sea pens) invariably bear distinct branches, often springing from a common stem. The branches must be separated so that water, bringing oxygen and nutrients, can reach the individual polyps (members of the colony) growing on them. At first glance, the Ediacaran “sea pens” look superficially like their modern counterparts in general shape, but they form a continuous, quilted structure, not a set of separated branches—and could therefore not operate like a modern soft coral colony. The Ediacaran “worms” are segmented and bilaterally symmetrical like their supposed modern descendants but many other creatures share the same symmetry—and such a basic and repeatable architecture need not imply close relationship. In other respects, the Ediacaran creatures are most unwormlike. They stretch up to a meter in length and remain flat as a pancake—more like films than the substantially thickened bodies of most modern segmented worms.
After exposing the differences between Ediacaran animals and their supposed modern counterparts, Seilacher examined the similarities that unite all Ediacaran forms. They share an architecture only rarely used by modern animals—and not by any living creature ever linked to an Ediacaran fossil. They look like ribbons, pancakes, and films, sometimes slightly “blown up” as air mattresses with a foliate or quilted structure.
The Ediacaran animals evolved before any creature had invented mineralized skeletons or external hard parts. Perhaps their unique
Bauplan
(to use the convenient German term for a basic scheme of organic architecture), records a pathway to large size that animals without supporting hard parts might follow—light and thin structures, woven together for added strength. In any case, and following a favorite theme of these essays for more than a decade, the Ediacaran fossils seem to represent one of two possible solutions—the one
not
followed by modern animals—to the basic structural problem of large size: the imposed decline of relative surface area since surfaces (growing as length squared) must increase more slowly than volumes (increasing as length cubed) as objects of similar shape get bigger. Since so many organic functions depend upon surfaces (respiration and feeding, to name just two) yet must serve the entire body’s volume, such a decline in relative surface cannot be tolerated for long.
Of the two possible solutions, nearly all large modern animals have retained their rounded or globular shapes but have evolved internal organs to increase surface areas—the rich branching of airways in our lung, and the complexly folded surface of our small intestine, for example. Another potential solution, followed rarely today but exploited by some large parasites, including tapeworms, permits large size without internal complexity by changing the body’s basic shape into something very thin—a ribbon or pancake—so that no internal space will be far from the external surface, the only site for respiration and absorption of food in the absence of internal organs. The Ediacaran animals, as a group, followed this second pathway to large size and therefore represent a coherent fauna strikingly different from any modern creature in basic design.
I might, were I inclined to search for progress in history, be gratified that life’s first “try” used the simpler of two solutions—a change in body shape rather than an invention of complex internal organs. However, the more important point remains that if Seilacher is right, the Ediacaran fauna represents a different, unique, and coherent experiment in organic architecture—not a set of precursors for modern animals. To emphasize this discontinuity, the first Paleozoic fauna with hard parts, the so-called Tommotian assemblage, is filled with tiny tubed, coiled, and cap-shaped creatures bearing precious little resemblance to Ediacaran forms. The ancestry of these later creatures may be recorded in indirect evidence for other Precambrian animals not included among the Ediacaran fossils. We have found abundant remains, in “trace fossils” of tubes for feeding and burrowing but, alas, no “body fossils” as yet, of animals with more conventional rounded shapes—a good source for later Tommotian descendants.
Seilacher ended his paper with a particularly arresting argument. We have, he pointed out, been searching with no success, and little hope, for complex extraterrestrial creatures, primarily because we wonder so powerfully what an independent experiment in the development of life might produce. What similarities would another “try” show with life on earth? How strong a constraint do the physics and chemistry of objects impose? How different could life be elsewhere? Our answers may lie in the concrete evidence of our own fossil record, and not in the abstract speculations of exobiology. Perhaps an independent experiment occurred right here on earth, expressing itself as the Ediacaran fauna, our first assemblage of multicellular animals.
Returning to the theme of mass extinctions, we used to argue that the first era boundary, between Precambrian and Paleozoic some 570 million years ago, presented a puzzling difference from all others because it recorded a profound radiation (the Cambrian explosion) but no previous extinction. But if the Ediacaran fauna, lying just below the base of the Paleozoic in strata throughout the world, represents a coherent and different experiment in life’s architecture, then a major extinction marks this initial boundary as well. The first strategy for mitigating mass extinction fails, and we trace little continuity across the opening and most profound boundary of life’s complex history.
Other papers at Indianapolis challenged the second strategy by arguing for a greater separation in effect and magnitude between mass extinctions and events of ordinary times. Some conclusions of previous years, already documented in these essays, have paved the way: (1) An asteroidal impact as the source, or at least the
coup de grâce
, of our terminal Cretaceous extinction (essay 25 in
Hen’s Teeth and Horse’s Toes
)—organisms, after all, can scarcely “prepare” for such a trigger. (2) David Raup’s estimate (essay 26 in
Hen’s Teeth and Horse’s Toes
) that a 50 percent extirpation of families, the counted figure for the Permian extinction, might translate to as much as 96 percent of all species (a removal of half the families implies an extinction of many more species since most species die without eliminating their families—a more inclusive category—while the death of a family must include all its species). For a removal so profound, we must seriously consider the possibility that entire groups will be lost for purely random reasons. (3) The calculation of Raup and Jack Sepkoski (essay 27 in
Hen’s Teeth and Horse’s Toes
) that major extinctions stand higher and more distinctly above the background level than previously recognized.