Authors: Sebastian Seung
Connectome (29 page)
Similarly, we might believe that memories could be read out from connectomes, if only there were a small success in this direction. Finding an entire human connectome still lies far in the future. For now, we have to work with partial connectomes derived from small pieces of brain. Perhaps we could pick a small chunk of human brain and attempt to read out memories from it. Or a piece of animal brain?
One thing is certain: Seeing the connectome will be just the first step. To read a book, you must do more than see the text. You must know the language in which the book is written, not to mention the letters of the alphabet and the spellings of words. In more technical terms, you must know how information is
encoded
in the marks on paper. Without knowledge of the code, a book is just a bunch of meaningless markings. Likewise, to read memories, we will have to do more than see connectomes. We will have to learn how to decode the information they contain.
Â
In what region of the human brain are we likely to find memories? Important clues come from the life of Henry Gustav Molaison, who died at a nursing home in Connecticut in 2008. During his lifetime he was known to the world as H.M., to protect his privacy. Many doctors and scientists studied H.M., who became one of the most famous neuropsychological cases since Broca's patient Tan.
In 1953, at the age of twenty-seven, H.M. underwent surgical treatment for severe epilepsy. The surgeon believed that H.M.'s seizures were originating in the medial temporal lobe (MTL), so he removed this region from both sides of H.M.'s brain. After surgery, H.M. appeared normal. His personality, intellect, motor skills, and sense of humor were intact. But there was an important and utterly incapacitating change. For the rest of his life, H.M. woke up every morning in a hospital room with no idea why he was there. He could not learn the names of the caretakers he saw every day. He could not name the president
or describe current events. In contrast, H.M. could still remember events from his life that had occurred before the surgery. The MTL seemed essential
for storing new memories but not for retaining old ones.
You probably remember that Itzhak Fried and his colleagues found the Jennifer Aniston and Halle Berry neurons in the MTL, showing that this region is involved in both perception and thought. Further experiments have explored its role in memory recall.
A patient was first shown many short video clips (five to ten seconds each) from cartoons, sitcoms, movies, and so on, while the activity of an MTL neuron was recorded. Later on, the patient was asked to freely recall the videos and report verbally whenever one came to mind. (During this second part of the experiment, no video was shown to the patient.)
One neuron spiked when the patient viewed a video of Tom Cruise but was much less active for other celebrities, places, and so on. Later on, the same neuron spiked whenever the patient reported recalling Tom Cruise but not when recalling other videos. Other neurons behaved similarly, being selectively activated by viewing or recalling one video but not others.
Perhaps the Tom Cruise neuron belongs to a cell assembly in the MTL. Perceiving or recalling Tom Cruise activates the cell assembly and hence the Tom Cruise neuron. If we want to read memories from connectomes, why don't we start by looking for cell assemblies in MTL? Unfortunately, the MTL is so large a region that finding its connectome is too difficult to be practical with our current technology.
We could narrow our search by focusing on the hippocampus, a part of the MTL thought to be important for storing new memories. In particular, the CA3 region of the hippocampus contains neurons that make synapses onto each other. Perhaps these connections allow groups of CA3 neurons
to form cell assemblies. But the human CA3
region is still rather large, so finding its connectome is currently out of reach. If we want to read memories, we had better find a smaller piece of brain to start with.
Â
H.M.'s amnesia only applied to
declarative
memory, which involves information that can be explicitly stated or “declared.” It includes autobiographical
events (“I broke my leg skiing last year”) as well as facts about the world (“Snow is white”). This is the most common meaning of the term
memory
.
There are also
nondeclarative
forms of memory, which involve information that is implicit rather than explicitly stated. These include motor skills and habits. H.M. could learn new motor skills, such as tracing a shape with a pencil while viewing his hand in a mirror. Based on his case and other types of evidence, neuroscientists have concluded that declarative and nondeclarative memory are distinct faculties, and perhaps served by different brain regions.
These two types of memory share some features, however. In his treatise
On Memory,
Aristotle compared recollection to movement: “Acts of recollection, as they occur in experience, are due to the fact that one movement has by nature another that succeeds it in regular order.” One can imagine that sequential memories, whether declarative or nondeclarative, are retained in the brain as synaptic chains. Perhaps the finger movements of a piano sonata played from memory are driven by sequential spiking of a synaptic chain somewhere in the pianist's brain.
It's difficult to study declarative memory in animals,
who can't tell us what they recall. But animals are perfectly capable of storing implicit memories. Why not try to read these from animal connectomes? I propose that we do this by searching for synaptic chains in the brains of birds.
Although birds are warm-blooded animals like us, they are more distant evolutionary cousins of ours than rodents. Since they don't nurse their young,
they're not classified as mammals. But mammals have no monopoly on intelligence. Despite the use of “birdbrain” as an insult, birds are actually quite smart. Mockingbirds and parrots excel at vocal mimicry, and crows can count and use tools. Because of these sophisticated behaviors, neuroscientists have been increasingly interested in our avian relatives.
Many study the zebra finch, a small Australian native that has spread throughout the world as a lovely pet. Males are adorned with orange cheeks and striking black and white patterns over the rest of their bodies. The male finch in Figure 44 is singing to the female, inviting her to mate. Males of other species also sing
to warn other males to keep out of their territory. All that twittering isn't meant for us, but it sounds beautiful just the same. Other birds too are popular pets because of their singing abilityâcanaries, for instance. Mozart kept a pet starling,
and taught it to trill a theme from the finale of one of his concertos. (Some claim the opposite, that the bird inspired his compositions.) Since birdsong makes use of pitch, rhythm, and repetition, some call it “nature's music.” Others compare it to language, as when the nineteenth-century master Percy Bysshe Shelley wrote of his art, “A poet is a nightingale who sits in darkness and sings to cheer its own solitude with sweet sounds.”
Â
 Â
Â
Â
Figure 44. Male zebra finch singing to female
Â
You might think that song is instinctive. Perhaps baby birds spring from the egg already knowing how to sing? No; those who have suffered through piano lessons need not be envious. The zebra finch does not acquire its talents effortlessly. Before starting to make sounds, a young male first hears his father's song. Later he starts to “babble,”
like a human baby making nonsense sounds. Over the next few months he practices singing tens of thousands of times, and ultimately learns to copy his father's song.
As a mature adult, a zebra finch sings essentially the same song every time. He does not improvise like a jazz pianist; he's more like a skater tracing compulsory figures on the ice. The song is said to be “crystallized.” The bird has stored a memory of its song and can recall it at will.
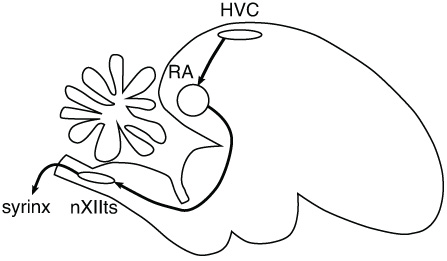
To produce sounds, birds use a vocal organ called the syrinx, which is like our larynx. Forcing air through the syrinx causes its walls to vibrate like a wind instrument. The pitch and other properties of the resulting sound are controlled by muscles around the syrinx,
which in turn receive instructions from the bird's brain. In the 1970s, Fernando Nottebohm identified the relevant regions of the brain, shown in the diagram in Figure 45. The names of the regions are long and complicated, so scientists simply use the abbreviations HVC, RA, and nXII.
Â
 Â
Â
Â
Figure 45. Song-producing regions of the bird's brain
Â
To understand the roles of the regions, let's compare this system to an artificial one for producing music. Perhaps you have a friend who is fanatic about high-end stereo equipment. Such audiophiles aren't satisfied with all-in-one systems; they like to have many separate components. In your friend's expensive stereo system, the compact disc player generates electrical signals, which travel to the preamplifier and then to the amplifier, and are finally transformed into sounds by the loudspeakers. In a bird's brain, electrical signals travel along an analogous pathway from HVC to RA to nXII, and are finally converted into sounds
by the syrinx. Every time the stereo plays Beethoven's Fifth Symphony, both the electrical signals in its components and the sounds from the loudspeaker are repeated in exactly the same sequence. Likewise, both the sounds from the syrinx and the spikes of neurons are repeated exactly the same way every time the bird sings.
Let's take a closer look at HVC. This region comes first in the song pathway, like the compact disc player of the stereo. Its name was originally “hyperstriatum ventrale, pars caudale,” or “HVc” for short. Later on, Nottebohm changed the name to “high vocal center,” abbreviated “HVC.” In 2005 a committee of neuroscientists decided that the letters stand for nothing.
(The situation is like that of the SAT, which once stood for “Scholastic Aptitude Test” and then for “Scholastic Assessment Test”; now its owner and developer, the College Board, offers no meaning at all.)
The name change came about because Harvey Karten, a specialist in brain structure and evolution, had convinced his colleagues that birds' brains are more similar to ours than previously thought. Neuroscientists had previously considered HVC analogous to the mammalian striatum, which is part of the basal ganglia, and believed that birds lack anything comparable to the neocortex. But Karten argued that a region called the dorsal ventricular ridge
functions like the neocortex. It contains a number of subregions that are believed important for the sophisticated bird behaviors mentioned above. One of these subregions is HVC.
Michale Fee and his collaborators
have measured spikes in HVC in live birds while they're singing. Some HVC neurons send their axons to RA, and they're the ones of interest here, as their signals travel along the song pathway. A zebra finch song consists of a few repetitions of a single motif. During a motif, which lasts 0.5 to 1 second, the neurons spike in a highly stereotyped sequence. In Figure 46 I've cartooned the spikes of three neurons. Each neuron waits until its moment in the motif comes, spikes for a few milliseconds, and then falls silent again. The time of spiking is precisely locked to a particular moment of the motif. This kind of sequential spiking is exactly what we'd expect from a synaptic chain.
Â
 Â
Â