Full House (16 page)
Darwin began by drawing a distinction between two kinds of "struggle" in his famous phrases—"struggle for existence" and "survival of the fittest." Struggle may take place directly against other organisms for limited resources (a type of competition called biotic), or against the rigors of the physical environment (called abiotic, or not involved with other living forms):
I should premise that I use the term Struggle for Existence in a large and metaphorical sense.... Two canine animals in a time of dearth, may be truly said to struggle with each other which shall get food and live. But a plant on the edge of a desert is said to struggle for life against the drought (1859, page 62).
Abiotic competition (the plant at the edge of the desert) cannot yield progress, for physical environments do not change in a persistent direction through time, and local adaptation can produce only a set of backings and forthings as lineages evolve first one way and then the other. But Darwin felt that biotic competition (two canine animals in a time of dearth) might yield progress—for if you are struggling with other members of your own species, rather than against a physical habitat, a more general biomechanical improvement transcending the particulars of any given environment—running faster, enduring longer, thinking better—might be your best option under natural selection. Thus, Darwin continued, if biotic competition is much more important than abiotic competition in the history of life, a general trend to progress might be defended.
But this argument for the prevalence of biotic competition will not suffice; another step is required. If environments are relatively empty—ei— ther because defeated forms can migrate somewhere else, or because losers can survive by switching to some other food or space in the same environment —then biomechanically inferior forms can continue to exist, and no ratchet to general progress will exist. But if ecologies are always chock-full of species, and losers have no place to go, then the victors in biotic competition will truly eliminate the vanquished—and the buildup of these successive eliminations might produce a trend to general progress. In fact, Darwin strongly advocated such a concept of nature’s plenitude—a notion that he tended to defend with a striking metaphor of the "wedge." Darwin depicts nature as a surface covered with wedges hammered into the ground and filling all space. A new species (depicted as a homeless wedge) can find a dwelling place only by discovering a tiny space between two existing wedges and hammering itself in by forcing another wedge out. In other words, each entry requires an expulsion
—
and biomechanical improvement might be the general key to successful wedging:
Nature may be compared to a surface covered with ten thousand sharp wedges ... representing different species, all packed closely together and all driven in by incessant blows ... sometimes a wedge ... driven deeply in forcing out others; with the jar and shock often transmitted far to other wedges in many lines of direction (from an 1856 manuscript, published by Stauffer, 1975).
Darwin then summed up his argument about biotic competition in a persistently full world by writing this and similar passages in the Origin of Species:
The inhabitants of each successive period in the world’s history have beaten their predecessors in the race for life, and are, in so far, higher in the scale of nature; and this may account for that vague yet ill-defined sentiment, felt by many paleontologists, that organization on the whole has progressed (1859, page 345).
I do not say that any obvious error pervades the logic of this argument, but we do need to inquire why Darwin bothered, and why the issue seemed important to him. Darwin had just devised an argument against progress—that the "bare bones mechanics" of natural selection yields only local adaptation, not general advance—and he had reveled in the radical character of this claim. Why, then, did he bother to smuggle progress back in through the rear door of a complex and dubious ecological argument about the predominance of biotic competition in a persistently full world? (Darwin surely recognized the shaky character of his necessary premise. He provided no clear rationale for biotic predominance—and Kropotkin and other critics would nail him on this point later. And the fossil record argued strongly against a persistently full world on a crucial issue that caused Darwin no end of trouble. Life’s history has been punctuated with several episodes of mass extinction; the largest, at the end of the Permian period, 250 million years ago, wiped out some 95 percent of the species of marine invertebrates. Clearly, habitats could not have been full after such episodes. Therefore, any buildup of progress between mass extinctions should be undone by the next dying. Darwin feared this argument greatly, and could extract himself only by claiming that mass extinctions were artifacts of an imperfect fossil record, an idea that can now be disproved with hard evidence for the triggering of at least one great dying by impact of an extraterrestrial body—the Cretaceous event that wiped out dinosaurs and gave us mammals a chance.)
I have no special insight into Darwin’s psyche, but I do feel that his strained and uncomfortable argument for progress arises from a conflict between two of his beings—the intellectual radical and the cultural conservative. The society that he loved, and that brought him such reward, had enshrined progress as its watchword and definition (I think of Herbert Spencer’s famous essay "Universal Progress, Its Law and Cause"). Darwin could not bear to fail his own world by denying its central premise. Yet his basic theory required just this opposition. So he forged an escape, and concocted a tenuous resolution by scaffolding a separate argument about ecology onto an edifice that could not support the required proposition by its own unique and different strength. But buildings with scaf folds look messy and incomplete—so why erect such a covering over a lovely structure that stands ever so well all by itself? I know no better illustration of the cultural power that progress holds over us than this story of Darwin’s own unresolved intellectual struggle, this tug-of-war between the logic of his theory and the needs of his society. If Darwin could not liberate himself from this deepest presupposition of our shared culture— even after inventing the theory with the key to this conceptual lock—then why should we be doing any better?
Fine. We may identify our assumption that evolution must entail progress as a cultural bias, and we may recognize that no good scientific argument for expecting progress exists, no more so in our own time than in Darwin’s day. We may also acknowledge that all standard attempts, including Darwin’s own, lie mired in social presupposition for the impetus, logical weakness for the argument, and factual inadequacy for the evidence.
And yet, undeniably (even for such curmudgeons as me), a basic fact of the history of life—the basic fact, one might well say—seems to cry out for progress as the central trend and defining feature of life’s history. The first fossil evidence of life, from rocks some 3.5 billion years in age, consists only of bacteria, the simplest forms that could be preserved in the geological record. Now we have oak trees, praying mantises, hippopotamuses, and people. How could anyone deny that such a history displays progress above anything else?
But every apparent certainty breeds subsequent doubt. Yes, peccaries, petunias, and poetry. But the earth remains chock-full of bacteria, and insects surely dominate among multicellular animals—with about a million described species versus only four thousand or so for mammals. If progress is so damned obvious, how shall this elusive notion be defined when ants wreck our picnics and bacteria take our lives? This very confusion permeates the fascinating colloquy between the Huxley and Darwin grand-children, as quoted at the beginning of this chapter. The modern Darwin asks the right questions, just as his grandfather did: How can "higher" be defined in an evolutionary world that produces a parasite for each supposed gain in progress? The modern Huxley gives a confused answer that unknowingly contains the germ of resolution: "I mean a higher degree of organization in general, as shown by the upper level attained." But to grasp the germ and unravel the confusion, we must reconceptualize the entire subject in a fundamental way—the same way that allowed us to resolve the paradox of 0.400 hitting; the same way that forms the subject of this entire book: viewing a history of change as the increase or contraction of variation in an entire system (a "full house"), rather than as a "thing" moving somewhere.
Claims for progress represent a quintessential example of conventional thinking about trends as entities on the move. From life’s infinite variety, we extract some "essential" measure like "average complexity" or "most complex creature"—and we then trace the supposed increase of this entity through time (as illustrated in the opening example of this book— see Figure 1). We label this trend to increase as "progress"—and we are locked into the view that such progress must be the defining thrust of the entire evolutionary process.
I shall, for the rest of Part Four, follow the same strategy of all my other examples by trying to view the variation in life’s complexity as primary and irreducible. I shall then trace the history of this variation through time. Only in this more adequate way can we acknowledge the obvious fact of "once only bacteria, but now petunias and people as well"—and still understand that no pervasive or predictable thrust toward progress permeates the history of life. We will, in short, learn the deeper reason why Darwin was right when he granted his radical intellect sway over his traditional social values.
13
A Preliminary Example at Smallest Scale, with Some Generalities on the Evolution of Body Size
In the case of 0.400 hitting, I spoke of a limit or "right wall" of human biomechanical possibility, and I illustrated the decrease in variation of batting averages as the full house of hitters moved toward this upper bound. In this section on complexity in the history of life, I shall present something close to a "mirror image" case—an increase in total variation by expansion away from a lower limit, or "left wall," of simplest conceivable form. The cases may seem quite different at first: improvement in baseball as decrease in variation by scrunching up against a right wall of maximal achievement versus increase of variation by spread away from a left wall of minimal complexity, misconstrued as an inevitable, overall march to progress in the history of life.
But a vital and deeper similarity unites the two examples—for both represent the same mode of correction for the same kind of error. In both cases, the error involves false portrayal of a complete system of variation by a single "thing" or entity construed as either the average or the best example within the system. Thus we tried to map the changing status of batting through time by tracing the history of the best conceived as a separable entity (0.400 hitters). Since this "thing" disappeared through time, we naturally assumed that the entire phenomenon—hitting in general—had gotten worse in some way. But proper consideration of the full house— the bell curve of batting averages for all regular players—shows that 0.400 hitting (properly viewed as the right tail of this bell curve, and not as a separable "thing") disappeared because variation decreased around a constant mean batting average. I then argued that we must interpret this shrinkage of variation as an indication of general improvement in play through time. In other words, by falsely isolating 0.400 hitting as a thing to be traced by itself, we got the whole story entirely backwards. The partial tale of the "thing" alone seemed to indicate degeneration of hitting; proper consideration of changes in the full variation showed that disappearance of 0.400 hitting represents improvement in general play.
We have traditionally made the same error—and must now make the same correction—in studying apparent trends to increasing complexity, or progress in the history of life. Again, we have abstracted the full and rich complexity of life’s variation as a "thing"—by taking either some measure of average complexity in a lineage or, more often, the particular case judged "best" (the most complex, the brainiest)—and we have then traced the history of this "thing" through time. Since our chosen "thing" has increased in complexity through time (once bacteria, then trilobites, now people), how could we possibly deny that progress marks the definition and central driving principle of evolution?
But I shall try to make the same correction in this part by arguing that we must consider the history of life’s complexity as a pattern of change for the full system of variation through time. Under this properly expanded view, we cannot regard progress as a central thrust and defining trend— for life began with a bacterial mode next to the left wall of minimal complexity; and now, nearly 4 billion years later, life retains the same mode in the same position. The most complex creature may increase in elaboration through time, but this tiny right tail of the full house scarcely qualifies as an essential definition for life as a whole. We cannot confuse a dribble at one end with the richness of an entirety—much as we may cherish this end by virtue of our own peculiar residence.
Before presenting the full argument for all of life, I must first explain why a dribble moving in one direction need not represent the directed thrust of causality within a system—but may actually arise as a consequence of entirely random movement among all items within the system. I will then demonstrate, in the next section, that apparent progress in the history of life arises by exactly the same artifact—and that, probably, no average tendency to progress in individual lineages exists at all.
I shall first illustrate the argument as an abstraction—using a classic pedagogical metaphor beloved by teachers of probability. Then I shall provide an intriguing actual case for a lineage of fossils with unusually good and complete data. Since we live in a fractal world of "self-similarity," where local and limited cases may have the same structure as examples at largest scale, I shall then argue that this particular case for the smallest of all fossils—single-celled creatures of the oceanic plankton—presents a structure and explanation identical with an appropriate account for the entire history of life. Since we can approach these largely unknown plankters without the strong biases that becloud our consideration of life’s full history, we can best move to the totality by grasping this self-similar example of oceanic unicells.
The overall directionality in certain kinds of random motion—an apparent paradox to many—can best be illustrated by a paradigm known as the "drunkard’s walk." A man staggers out of a bar dead drunk. He stands on the sidewalk in front of the bar, with the wall of the bar on one side and the gutter on the other. If he reaches the gutter, he falls down into a stupor and the sequence ends. Let’s say that the sidewalk is thirty feet wide, and that our drunkard is staggering at random with an average of five feet in either direction for each stagger. (See Figure 21 for an illustration of this paradigm); for simplicity’s sake—since this is an abstract model and not the real world—we will say that the drunkard staggers in a single line only, either toward the wall or toward the gutter. He does not move at right angles along the sidewalk parallel to the wall and gutter.
Where will the drunkard end up if we let him stagger long enough and entirely at random? He will finish in the gutter-absolutely every time, and for the following reason: Each stagger goes in either direction with 50 percent probability. The bar wall at one side is a "reflecting boundary."
8
If the drunkard hits the wall, he just stays there until a subsequent stagger propels him in the other direction. In other words, only one direction of movement remains open for continuous advance—toward the gutter. We can even calculate the average amount of time required to reach the gutter. (Many readers will have recognized this paradigm as just another way of illustrating a preferred result in coin tossing. Falling into the gutter on one unreversed trajectory, after beginning at the wall, has the same probability as flipping six heads in a row [one chance in sixty-four]-five feet with each stagger, to reach the gutter in thirty feet. Start in any other position, and probabilities change accordingly. For example, once the drunkard stands in the middle, fifteen feet from the wall, then three staggers in the same direction [one chance in eight for a single trajectory] put him into the gutter. Each stagger is independent of all others, so previous histories don’t count, and you need to know only the initial position to make the calculation.)
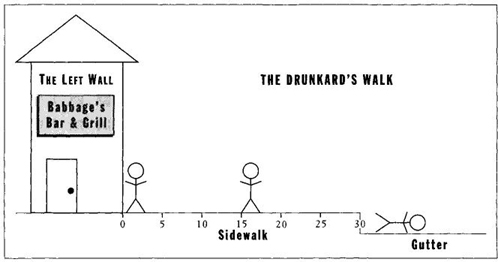
FIGURE 21 The drunkard’s walk.
I bring up this old example to illustrate but one salient point: In a system of linear motion structurally constrained by a wall at one end, random movement, with no preferred directionality whatever, will inevitably propel the average position away from a starting point at the wall. The drunkard falls into the gutter every time, but his motion includes no trend whatever toward this form of perdition. Similarly, some average or extreme measure of life might move in a particular direction even if no evolutionary advantage, and no inherent trend, favor that pathway.
Turning to a similar example in the history of life, Foraminifera are single-celled protozoans that secrete a skeleton around or within their protoplasm, and are therefore extremely common in the fossil record. (In fact, since they tend to be so abundant—ubiquitous in many marine sediments—they serve as some of the best markers for tracing time and environment in the geological record. Although most of the public never comes in contact with "forams"—as we in the trade call them for short— their study absorbs the lives of a large fraction of professional paleontologists.) Most marine forams live in bottom sediments, and are called benthic. A few species float in open waters near the oceanic surface, and are called planktonic. These planktonic forams are especially important in dating sediments, and in reconstructing former environments and movements of water masses, during the Cenozoic Era (the past 65 million years, since the extinction of dinosaurs). As a result of their mobility, planktonic species live over large areas of the globe, and are therefore particularly valuable in permitting comparison of sediments from widely separated places (most benthics have much more limited ranges, and consequently less utility).
The basic outline of the evolutionary history of modern planktonic forams has been well known for a long time. They arose in the Cretaceous (the last period of the Mesozoic Era, when dinosaurs dominated terrestrial ecosystems), and they remain vigorously alive today. Their evolution has been interrupted by two episodes of mass extinction, when most species died and only a few survived to continue the lineages: once at the end of the Cretaceous (one of the five great mass dyings in the history of life— this is the event that triggered the death of dinosaurs and almost surely involved the impact of a large extraterrestrial object as a fundamental cause); and again during the largest episode of extinction within the Cenozoic Era. Thus, the evolution of planktonic forams is a drama in three largely independent acts (linked by a few transitions): the Cretaceous for Act One, the earlier Cenozoic (called Paleogene) for Act Two, and the later Cenozoic (called Neogene) for Act Three.
Traditional wisdom, and any textbook, will tell you that each of the three acts follows the same pattern, thus making the entire story so famous in professional circles—for paleontologists crave independent repetition as a test for predictable results (the closest a historical scientist can come to the experimental ideal of replication under identical conditions in a laboratory). The founding lineages for each of the three radiations were small in body size—and size then increased (or so we are told) during each of the three evolutionary diversifications. If an identical result occurs in each of three episodes, then we are probably witnessing an evolutionary generality. In fact, paleontologists treasure this case as our best illustration of the one decent phylogenetic "rule" that the fossil record seems to affirm with copious evidence.
The attempt to establish such "rules," or generalities in evolution during geological time, absorbed much attention in generations past. But the strategy largely fizzled because few proposed "rules" survived the weight of accumulated exceptions in our complex and contingent world of evolutionary change. The one generality that survived, and that seems to hold firm as more evidence accumulates, is known as "Cope’s Rule" (after the brilliant and contentious nineteenth-century American vertebrate paleontologist)—the observation that most lineages tend to increase in body size during their evolutionary history. (Like all evolutionary generalizations, "Cope’s Rule" identifies a predominant relative frequency, not an absolute statement. Many lineages decrease in size. An increase of size in 70 percent of lineages, when we think that a random world should yield half and half, is more than enough for a "rule" in our trade.)
The evidence, as usually presented, certainly seems to support Cope’s Rule for plank
t
onic forams. Figure 22 shows the increase in body size over time for both the largest species and the average of all species during Act One of the Cretaceous Period (figures for Acts Two and Three show the same pattern). I shall not deny this evidence of increase in each act for the largest or average species. But this book is dedicated to providing an enlarged perspective—offering a different and often opposite interpretation—for this very situation of "trends" myopically depicted as "things" moving somewhere rather than as changes in variation of entire systems ("full houses").
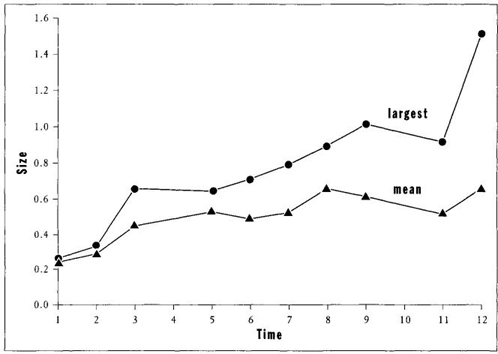
FIGURE 22 Inappropriate use of mean or extreme values to specify a trend of increasing size where none exists when the full range of variation is properly considered.