Full House (8 page)
W. D. Matthew, author of the most famous icon for the equine ladder (Figure 9), fell into the same error because his designation of one pathway as a main line forced him to interpret all others as diversions of lesser value. Matthew (1926, page 164) called his ladder "the direct line of succession," and added that "there are also a number of side branches, more or less closely related." But Matthew then imposed a brand of near inde
-
cency upon his previous charge of mere laterality, as he described (1926, page 167) "a number of side branches leading up ... to aberrant specialized Equidae now extinct." But in what way are these extinct lineages more specialized than a modern horse, or in any sense more peculiar? Their phyletic death sets the only possible rationale for a designation of aberrancy, but more than 99 percent of all species that ever lived are now extinct—and disappearance is not the biological equivalent of a scarlet letter.
I have thus far presented the case of horses only as a general argument about bushiness versus linearity. I do not deny the factuality of the conventional pathway and its trends in size, teeth, and toes, but I do wish to demonstrate what a distorted—even backward—view this little piece provides when we depict Hyracotherium → Equus as the essence of the history of horses, and then ignore the variation supplied by a myriad of other pathways in the full house of the equine bush. Three categories of detail should cement the importance of the opposite perspective gained by considering the changing spread of variation through time—horses as a declining lineage within a failing larger group.
1. The evolutionary tree of horses is copiously bushy throughout; no geological segment of equine history can be construed as featuring a wide main line with dribbling side branches. Bruce MacFadden of the Florida Museum of Natural History, our leading modern expert on the paleobiology of horses, recently published a simplified picture of the equine tree (reproduced here as Figure 10). Consider the last 20 million years from the key transition of browsing to grazing, up to modern times. We note only copious branching bushiness, and nothing that anyone could identify as a central thrust amid the diversity. MacFadden couldn’t even begin to depict all the complex branching in his single diagram, so he expanded a key portion (as indicated by the square that he drew over Figure 10), and presented a fuller account of this 7-million-year interval (reproduced here as Figure 11). In North America alone, between 15 and 18 million years ago, at least nineteen species originated by branching. By 15 million years ago, sixteen contemporaneous grazing species inhabited North America (while several older lineages of browsing horses also lived in America and the Old World). This diversity hardly changed during the next 7 million years "as extinctions balanced originations, resulting in a steady-state diversity pattern" (MacFadden, 1988, page 2). North American diversity then declined rapidly, and the entire bush of horses eventually died out in the New World. (Remember how Cortez’s horses terrified the Aztecs, who had never seen the beasts that had originated on their continent and then become extinct. Eurasia was an outpost of equine survival, not a center for an expanding trend.)
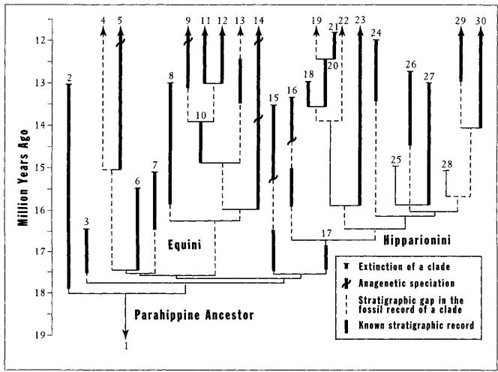
FIGURE 11 So much branching occurs in the evolution of horses during the Middle Miocene that MacFadden, as shown in Figure 10, could not include all the individual lineages. This figure is an enlargement of the box shown in Figure 10. Note how many branching events occurred during this relatively short interval.
Two points stand out in this epitome for the last third of equine history. First we note a primary signal of branching, branching, and more branching. Where, in this forest, could anyone identify a main trunk? The bush has many tips, though all but one, the genus Equus, are extinct. Each tip can be connected to a last common ancestor by a labyrinthine route, but no paths are straight, and all lead back by sidestepping from one event of branching speciation to another, and not by descent down a ladder of continuous change. If you venture an argument that the pathway to modern Equus should be viewed as a main line because the genus still lives and once spread (by its own devices, and not by human transport) over all major continents, I reply that Equus died out over most of its range, including the North American fatherland, and that all modern species derive from Old World remnants. Second, I think that any unbiased observer must identify decline as the major feature of equine evolution during the last 10 million years—the very period when traditional ladder models proclaim perfection and fine-tuning of the distinctive trend to a single hoofed toe, with side toes reduced to vestigial splints. An average of sixteen contemporaneous species lived in North America alone from about 15 million to about 8 million years ago—until, to invoke Agatha Christie’s famous image, they died one by one—and then there were none.
Rearguard defenders of the ladder might reply that I have been discussing only the last (and admittedly bushy) third of equine evolution. What about the first 40 million years, shown as tolerably linear even on MacFadden’s arborescent picture (Figure 10)? This earlier period has been the chief domain for friends of linearity. Even G. G. Simpson, who began the transition to bushy thinking in his wonderful 1951 book, Horses, and who drew the first famous arborescent diagram of equine phylogeny (a less bushy ancestor of MacFadden’s version, reproduced here), defended the basic linearity of this earlier record. "The line from Eohippus [Hyracotherium] to Hypohippus, " he wrote (1951, page 215), "exemplifies a fairly continuous phyletic evolution." Simpson especially emphasized the supposedly gradual and continuous transformation from Mesohippus to Miohippus near the top of this sequence (see Figure 10 for all names and times):
The more progressive horses of the middle Oligocene ... are placed by convention in a separate genus, Miohippus. In fact Mesohippus and Miohippus intergrade so perfectly and the differences between them are so slight and variable that even experts find it difficult, at times nearly impossible, to distinguish them clearly.
The enormous increase in fossil evidence since Simpson’s time has allowed paleontologists Don Prothero and Neil Shubin (1989) to falsify this view, and to introduce extensive bushiness into this last stronghold of the ladder, as predicted by the theory of punctuated equilibrium (see Eldredge and Gould, 1972; Gould and Eldredge, 1993). Prothero and Shubin made four major discoveries in this early segment of equine history that Simpson had designated as the strongest case for a gradual sequence of linear transformation—the transition from Mesohippus to Miohippus.
First, the two genera can be sharply distinguished by features of the footbones, previously undiscovered. Mesohippus does not grade insensibly into Miohippus. (Previous claims had been based on teeth, the best preserved parts of mammalian skeletons. The genera cannot be distinguished on dental evidence—the major criterion available to Simpson.)
Second, Mesohippus does not evolve to Miohippus by insensible degrees of gradual transition. Rather, Miohippus arises by branching from a Mesohippus stock that continues to survive long afterward. The two genera overlap in time by at least 4 million years.
Third, each genus is itself a bush of several related species, not a rung on a ladder. These species often lived and interacted in the same area at the same time. One set of strata in Wyoming, for example, has yielded three species of Mesohippus and two of Miohippus, all contemporaries.
Fourth, the species of these bushes tend to arise with geological suddenness, and then to persist with little change for long periods. Evolutionary change occurs at the branch points themselves, and trends are not continuous marches up ladders, but concatenations of increments achieved at nodes of branching on evolutionary bushes. Prothero and Shubin write,
This is contrary to the widely held myth about horse species as gradualistically varying parts of a continuum, with no real distinctions between species. Throughout the history of horses, the species are well-marked and static over millions of years. At high resolution, the gradualistic picture of horse evolution becomes a complex bush of overlapping, closely related species.
In other words, bushiness now pervades the entire phylogeny of horses.
2. Plausible alternative histories would have yielded a very different and not nearly so attractive story. The substitution of bushes for ladders certainly calls into question, but does not necessarily falsify, the conventional lockstep view of transitions to fewer toes, larger bodies, and higher-crowned teeth. After all, older branches of a bush need not endure for long, and their early removal would leave no ancient vestiges to compromise a trend by persistent variation. If all the early branches die, and all the later twigs bear "progressive" features, then the tree becomes "modernized" throughout—and we may fairly talk of a pervasive trend. If all small horses die early, if no three-toed horses survive into the regime of one-toed Equus, then we may justly speak of general trends to increased size and a single hoof—and the old marching order from Hyracotherium to Equus might be defended as a fair epitome of real directionality (while still subject to criticism for neglecting the equally important pattern of waxing and waning diversity). In such a world, the objections that I have raised would be carping and trivial. Yes, we could still emphasize that many pathways run through the bush, and that Hyracotherium to Equus marks only one lineage—but if all pathways pass through the same sequence to larger size and fewer toes, then any one will show the genuine trend, and we shouldn’t be too critical if convention favors one case over all others.
This last-ditch defense of equine progress cannot be sustained. The conventional trends are by no means pervasive (though their relative frequency does increase through the bush, albeit in a fitful way). Several late lineages negate the most prominent trends, and a different outcome for the history of horses—perfectly plausible in our world of contingency (see Gould, 1989)—would have compelled a radically altered tale.
Consider just one arresting scenario. Contrary to the usual view that horses increase inexorably in body size, MacFadden (1988) studied all ancestral-descendant pairs of species that he could identify with confidence on the equine bush. Of twenty-four such pairs, he found that five, or more than 20 percent, showed a decrease in size. Dwarfing has been a common and persistent phenomenon, repeated throughout the history of horses. Even the first genus, Hyracotherium, included periods of size decrease during its geological history (see Gingerich, 1981).
The most recent, and most profound, trend to dwarfing occurred in a North American genus appropriately named Nannippus (or dwarfed horse). Simpson writes of this remarkable genus (1951, page 140): "Some of the late specimens were miniatures no higher than a small Shetland pony and considerably more slender. These graceful creatures had long, thin legs and feet, and the general form probably suggested a small gazelle more than an ordinary horse."
Now suppose that Nannippus had survived as the only living member of the Equidae, and Equus had died or never arisen. How would we then tell the story of horses in our biased mode of running steamrollers over one pathway through the bush and calling the resulting line canonical? I hear you crying "foul." You say that Nannippus was a funny little side branch and Equus a powerful main line—so I must be playing verbal games with a story that could never have occurred. Not so; my tale is plausible, but just unrealized. Nannippus showed substantial geographic breadth and geological depth. The genus lived in the United States and Central America, arose more than 10 million years ago, and failed to survive by only a whisker, becoming extinct only about 2 million years ago (MacFadden and Waldrop, 1980). Four species have been described (MacFadden, 1984), and their range of some 8 million years greatly exceeds the longevity of Equus (see Figure 11). If you say that Equus had a greater chance because this modern genus spread from an American homeland into Eurasia and Africa, while Nannippus never colonized the Old World, I reply that Equus became entirely extinct throughout its hemisphere of origin, and therefore only survived by a whisker itself. Suppose that Nannippus had migrated and Equus stayed at home?
What would be left of our vaunted horse story if Nannippus had survived, and Equus died ? We wouldn’t be advertising any drive to increased size because Nannippus, though a dwarfed descendant of larger ancestors, isn’t much bigger than the original Hyracotherium. We wouldn’t be getting very excited about reduction in toes either, because Nannippus still sported three on each foot (though the side toes were reduced), whereas the original Hyracotherium had four toes on the front feet and three behind (not five on each limb, as commonly misconstrued). We would be left, in fact, only with the trend to increased crown height of the molar teeth— and here we could gloat, because Nannippus chewed with the relatively tallest teeth of any horse in history, including modern Equus. But then, tooth height has never provided much of a draw for museums or textbook diagrams, and the conventional story rests upon reduction of toes and growth of body. In short, if Nannippus had survived and Equus died, we wouldn’t be telling any famous story about horses at all. The equine bush would become just another anonymous part of the rich mammalian record, known to specialists and unadvertised to the public. Yet nothing would be different but the substitution of one twig for another at the very end of a rich history.