Power, Sex, Suicide: Mitochondria and the Meaning of Life (7 page)
Read Power, Sex, Suicide: Mitochondria and the Meaning of Life Online
Authors: Nick Lane
Tags: #Science, #General
The point I want to make about methanogens is that they were the losers in the race through a bottleneck, yet nonetheless survived in niche environments. Similarly, on a larger scale, it is rare for the loser to disappear completely, or for the latecomers never to gain at least a precarious foothold. The fact that flight had already evolved among birds did not preclude its later evolution in bats, which became the most numerous mammalian species. The evolution of plants did not lead to the disappearance of algae, or indeed the evolution of vascular plants to the disappearance of mosses. Even mass extinctions rarely lead to the disappearance of whole classes. If the dinosaurs disappeared, the reptiles are nevertheless still among us, despite stiff competition from birds and mammals. It seems to me that the only bottleneck in evolution comparable to that which de Duve postulates for the eukaryotic cell is the origin of life itself, which may have happened once, or perhaps numerous times with only one form ultimately surviving—in which case this, too, was a bottleneck. Perhaps, but this is not a good example, for we simply don’t know. All we can say for sure is that all life living today ultimately shares the same ancestor, and so sprang from the same progenitor. Incidentally, this rules out the view, expressed by some, that our planet has been populated by successive waves of invasions from outer space—such a view is not compatible with the deep biochemical relatedness of all known life on earth.
If the origin of the eukaryotic cell was not a bottleneck, then it was probably a genuinely unlikely sequence of events, for it happened only once. Speaking as a multicellular eukaryote, I might be biased, but I do not believe that bacteria will ever ascend the smooth ramp to sentience, or anywhere much beyond slime, here or anywhere else in the universe. No, the secret of complex life lies in the chimeric nature of the eukaryotic cell—a hopeful monster, born in an improbable merger 2000 million years ago, an event still frozen in our innermost constitution and dominating our lives today.
Richard Goldschmidt first advanced the concept of a hopeful monster in 1940—the year that Oswald Avery showed that the genes are composed of DNA. Goldschmidt’s name has since been derided by some writers, and held up as an
anti-Darwinian hero by others. He deserves neither epitaph as his theory is neither impossible nor anti-Darwinian. Goldschmidt argued that the gradual accumulation of small genetic changes,
mutations
, was important, but could only account for the variation within a species: it wasn’t a sufficiently powerful source of evolutionary novelty to explain the origin of a new species. Goldschmidt believed that the big genetic differences between species could not be mounted by a succession of tiny mutations, but required more profound ‘macro-mutations’—monster leaps across ‘genetic space’, which is to say the gulf between two different genetic sequences (the number of changes required to get from one to another). He appreciated that random macro-mutations, sudden large changes in gene sequence, were far more likely to produce an unworkable mutant, and so he christened his one-in-a-million success a ‘hopeful monster’. For Goldschmidt, a hopeful monster was the lucky outcome of a large and sudden genetic change, rather than a succession of tiny mutations—the kind of thing an archetypal mad scientist might produce in the laboratory after a lifetime of dedicated and deranging failure. With our modern understanding of genetics, we now know that macro-mutations don’t account for speciation, at least not in multicellular creatures (though they may in bacteria, as argued by Lynn Margulis). However, it seems to me that the fusion of two whole genomes to create the first eukaryotic cell is better viewed as a macro-mutation to create a ‘hopeful monster’ than purely as a succession of small genetic changes.
So what kind of a monster was the first eukaryote, and why was its origin so improbable? To understand the answers, we need to think first about the nature of eukaryotic cells, and the many striking ways in which they differ from bacteria. We have already touched on this in the Introduction; here, we need to focus on the magnitude of the differences, the wide yawn of the chasm.
Compared with bacteria, most eukaryotic cells are enormous. Bacteria are rarely larger than a few thousandths of a millimetre (a few microns) or so in length. In contrast, although some eukaryotes, known as the
pico-eukaryotes
, are of bacterial size, the majority are ten to a hundred times those dimensions, giving them a cell volume about 10 000 to 100 000 times that of bacteria.
Size is not the only thing that matters. The cardinal feature of eukaryotes, from which their Greek name derives, is the possession of a ‘true’ nucleus. This nucleus is typically a spherical, dense mass of DNA (the genetic matter) wrapped up in proteins and enveloped in a double membrane. Here, already, are three big differences with bacteria. First, the bacteria lack a nucleus at all, or else have a primitive version that is not enclosed by a membrane. For this
reason bacteria are also termed ‘prokaryotes’, from the Greek ‘before the nucleus’. While this is potentially a prejudgement—some researchers argue that cells with a nucleus are just as ancient as those without—most specialists agree that prokaryotes are well named: they really did evolve before cells with a nucleus (the eukaryotes).
The second big difference between bacteria and eukaryotes is the size of their genomes as a whole—the total number of genes. Bacteria generally have far less DNA than even simple single-celled eukaryotes such as yeast. This difference can be measured either in terms of the total number of genes—usually adding up to hundreds or thousands—or the total DNA content. This latter value is known as the C-value, and is measured in ‘letters’ of DNA. It includes not only the genes, but also the stretches of so-called
non-coding
DNA—DNA that does not code for proteins, and so can’t really be called ‘genes’. The differences in both the number of genes and the C-value are revealing. Single-celled eukaryotes like yeasts have several times as many genes as most bacteria, whereas humans have perhaps twenty times as many. The difference in the C-value, or total DNA content, is even more striking, as eukaryotes contain far more non-coding DNA than bacteria. The total DNA content of eukaryotes spans an extraordinary five orders of magnitude. The genome of a large amoeba,
Amoeba dubia
, is more than 200 000 times larger than that of the tiny eukaryotic cell,
Encephalitozoon cuniculi
. This enormous range is unrelated to complexity, or the total number of genes.
Amoebae dubia
actually has 200 times more DNA than do humans, even though it has far fewer genes and is obviously less complex. This odd discrepancy is known as the C-value paradox. Whether all this non-coding DNA has any evolutionary purpose is debated. Some of it certainly does, but a large part remains puzzling, and it is hard to see why an amoeba should need so much (we will return to this in
Part 4
). Nonetheless, it is a fact, requiring an explanation, that eukaryotes generally have orders of magnitude more DNA than prokaryotes. This is not without a cost. The energy required to copy all this extra DNA, and to ensure it is copied faithfully, affects the rate and circumstances of cell division, with implications that we will explore later.
The third big difference lies in the packing and organization of DNA. As we noted in the Introduction, most bacteria possess a single circular chromosome. This is anchored to the cell wall, but otherwise floats freely around the cell, ready for quick replication. Bacteria also carry genetic ‘loose change’ in the form of tiny rings of DNA called plasmids, which replicate independently and can be passed from one bacterium to another. The daily exchange of loose plasmids in this way is equivalent to shopping with loose change, and explains how the genes for drug resistance spread so quickly in a population of bacteria—just as a coin may find itself in twenty different pockets in a day. Returning to their main gene bank, few bacteria wrap their main chromosome in proteins—rather,
their genes are ‘naked’, making them easily accessible—a current account rather than a savings account. Bacterial genes tend to be ordered in groups that serve a similar purpose, and act as a functional unit, which are known as
operons
. In contrast, eukaryotic genes give no semblance of order. Eukaryotic cells possess quite a number of disparate, straight chromosomes, which are usually doubled up to give pairs of equivalent chromosomes, such as the 23 pairs of chromosomes found in humans. In eukaryotes, the genes are strung along these chromosomes in virtually a random order, and to make matters worse they are often fragmented into short sections with long stretches of non-coding DNA breaking up the flow. To build a protein, a great tract of DNA often needs to be read off, before it is spliced up and melded together to form a coherent transcript that codes for the protein.
Eukaryotic genes are not just randomized and fragmented, they are also tricky to get at. The chromosomes are tightly wrapped in proteins called his-tones, which block access to the genes. When the genes are being replicated during cell division, or copied to make transcripts for building proteins, the configuration of the histones must be altered to allow access to the DNA itself. This in turn has to be controlled by proteins called transcription factors.
Altogether, the organization of the eukaryotic genome is a complicated business that fills library after library with footnotes. We’ll come to another aspect of this complicated set-up in
Part 5
(sex, which is not found in bacteria). For now, though, the most important take-home point is that there is an energetic cost to all of this complexity. Where bacteria are almost always ruthlessly streamlined and efficient, most eukaryotes are lumbering and labyrinthine.
Outside the nucleus, eukaryotic cells are also very different to bacteria. Eukaryotic cells have been described as cells with ‘things inside’ (
Figure 2
). Most of the things inside are membrane structures, made of a vanishingly thin sandwich of fatty molecules called lipids. The membranes form into vesicles, tubes, cisterns, and stacks, enclosing spaces—closets—that are physically separated, by the lipid barrier, from the watery cytosol. Different membrane systems are specialized for various tasks, such as building the components of the cell, or breaking down food to generate energy, or for transport, storage, and degradation. Interestingly, for all their variety of size and shape, most of the eukaryotic closets are variations on the simple vesicle: some are elongated and flattened, others are tubular, and some are simply bubbles. The most unexpected is the nuclear membrane, which looks like a continuous double membrane enclosing the nucleus, but is in fact a series of large flattened vesicles welded together, and rather surprisingly, continuous with other membrane compartments in the rest of the cell. The nuclear membrane is therefore distinct in structure from the external membranes of any cell, which are always a continuous single (or double) layer.
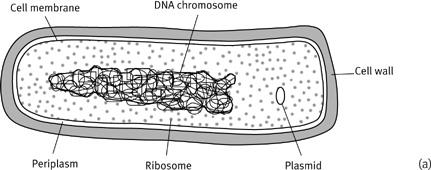
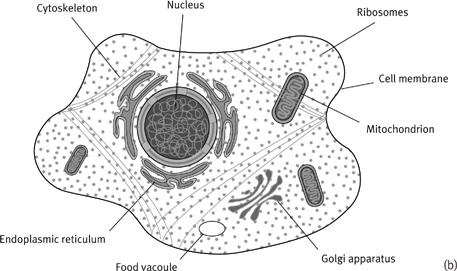
2
Schematic illustrations of a bacterial cell (a) compared with a eukaryotic cell (b). The illustrations are not drawn to scale; bacteria are about the same size as the mitochondria in (b). The membrane structures are in fact sparsely depicted in the eukaryotic cell, for clarity, and in reality the differences in internal structure are even more marked. Bacteria are remarkable for their inscrutability, even by electron microscopy.
Then there are the tiny organs within cells, the so-called organelles, such as the mitochondria and the chloroplasts in plants and algae. The chloroplasts are worth a special mention. They are responsible for photosynthesis, the process by which solar energy is converted into the currency of biological molecules, which possess their own chemical energy. Like the mitochondria, the chloroplasts
derive from bacteria, in this case the
cyanobacteria
, the only group of bacteria capable of true photosynthesis (to generate oxygen). It is notable that both mitochondria and chloroplasts were once free-living bacteria, and still retain a number of partially independent traits, including a contingent of their own genes. Both are involved in energy generation for their host cells. Both these organelles are tangibly different from the other membrane systems of eukaryotic cells, and these differences set them apart. Like the nucleus, the mitochondria and chloroplasts are enclosed in a double membrane, but unlike the nucleus their membranes form a true continuous barrier. Along with their own DNA, their own ribosomes and protein assembly, and their semi-autonomous manner of division, the double membranes of the mitochondria and chloroplasts are among the features that point an incriminating finger at their bacterial ancestry.