Regenesis (14 page)
Authors: George M. Church
Here's how it works. To modify a genome, MAGE uses the smallest bits of DNA that are unique in the genome to be changed. The smallest unique pieces are theoretically around 12-mers for 5 million base pair bacterial genomes, since the number of ways that you can arrange four bases (A, C, G, T) into words twelve base pairs long is 4
12
= 16 million. In practice, the optimum seems to be 90-mers. We get these 90-mers into cells by means of a short, 2,500-volt electrical pulse. We chemically armor the ends of the 90-mer against enzyme damage in the cell, coat them with a protein that enhances pairing, and then let these pieces sneak into the gaps wherever
the genome is replicating. Among other tricks, we use multiple 90-mers and multiple cycles. This collection of lab shortcuts and other techniques raises the chances of getting the desired genomic mutations from practically infinitesimal (one in a million) to very common (several per hour per cell). So, instead of assembling small DNAs into a huge piece of DNA, at great cost and with multiple errors, we use many small bits in parallel, a process that allows us to test several combinations for the purpose of seeing which work well and which are deleterious to the cell.
The other technique that greatly enables MAGE is selection, which is of course the key mechanism propelling Darwinian evolution. Selection in this context means rapidly separating out cells (or molecules) that have the more extreme values of the desired properties from all the rest. “Give me a fulcrum and I will move the world,” from Archimedes, gets updated in the synthetic genomics lab to, “Give me a selection, and I will change the world.”
We can easily select for leaving behind babies, since that is the best-tested selection paradigm of all time (tested by 10
27
organisms for 3 billion years each). The players who leave behind the most surviving grandchildren tend to dominate subsequent history. This is of course a selection for all of the processes required for replication, not just the properties that we want to enhance. But frequently human engineers want to select for some property other than replicationâfor example, for overproducing milk in cows or producing insulin in bacteria, or artemisinin (an anti-malarial precursor) in yeast.
Given that we want to engineer a whole genome, which is better? (1) Synthesize all of it, all at once, with possibly hundreds of design flaws to debug simultaneously? Or (2) make targeted changes combinatorially, dividing and conquering each bug swiftly and in parallel (and harnessing lab evolution). The Church lab voted strongly for (2).
Summing up, genome engineering already embraces much more than just genome copying, and much more than even the mature engineering disciplines (i.e., design and fabrication). Genome engineering has evolution on its sideânot slow evolution but intelligently designed, fast evolution to create a range of special-purpose organisms that can produce desired substances.
So now let's suppose that we have created entire new genomes, and perhaps even some new single-cell organisms. The next step, obviously, is multicellularity.
One of the first signs of multicellularity, even before cells clump and differentiate, is sex. Exchanging genetic material allows for highly parallel discovery of new genomic sections. For example, if it takes a day for a bacterial cell to replicate in the wild, it will take thirty days for a mutant cell to replace a population of about a billion cells, and it will take 100,000 years to change 20 percent of a bacterial genome (a working definition of a new species). This assumes that selection pressure is fairly steady in one direction and that there is a constant stream of beneficial mutations. In practice a significant fraction of the evolutionary steps toward that 20 percent are neutral, neither enhancing nor harming the organism, and the number of setbacks are such that a million years is not unusual for the full appearance of new species. If new favorable mutations can combine by means of sex, then the time spent sweeping away the competing population is saved (keeping in mind that sex can separate favorable alternative forms of a gene as well as bring them together).
To drive all this toward its limits, we are using a second lab techniqueâconjugative assembly genome engineering (CAGE)âselectively mating two bacterial strains in order to combine parts of their genomes. If MAGE is a means of breaking up a long piece of DNA and making small changes in each part, CAGE is a means of putting the changed parts together again. This allows us to divide and conquer the
E. coli
genome, for example, into thirty-two pieces. We first apply the MAGE technique, focusing on only 140 kbp in each of thirty-two separate strains in parallel. We then mate strain 1 and 2, bringing together adjacent sectors in the genomes of the strains. Similar matings bring together sixteen even-numbered sectors with the adjacent sixteen odd-numbered sectors, thereby reducing the number of strains by a factor of two. Then we repeat the process to get down to eight strains, then four, then two, and then finally one bacterial strain.
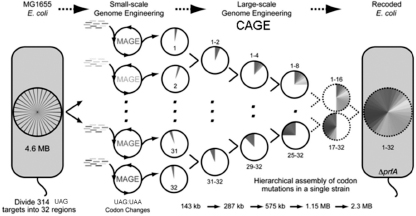
Figure 3.3
Divide and conquer with MAGE and CAGE. For example, to change all UAG stop codon targets (322 total in the
E. coli
genome) into the functionally identical codon UAA, we divide this task into thirty-two sectors of the genome pie (far left) and design thirty-two pools of ten oligos each (second column) that can change ten codons in each sector (third column). By selective mating we merge adjacent pairs of sectors (e.g., 1 and 2) resulting in sixteen bacterial strains, each with a double-size sector (fourth column) in the genome of that strain. This process of merging adjacent sectors continues to eight, then four, then two (columns 5 to 7); the final mating results in one strain that has all of the UAG codons changed to UAA, as was planned.
A second advantage of multicellularity is cross-feeding, where one cell type can focus on one type of food chemistry and can barter its specialty food for a different food from an adjacent cell. You can divide your genome over two or more cell types and hence save space, modularize and parallelize selection, and quickly benefit from sharing the multitude of tasks.
A third advantage of multicellularity is shape selection. Single cells have some diversity in morphology, but their limit becomes rapidly evident. For example, making a cell that is as hard as a tooth doesn't automatically confer chewing ability, since that requires structures larger and more complex than a cell. How to get from the straight (one-dimensional) DNA to complex three-dimensional dynamic shapes is still mysterious. But, as always, synthesis offers us both a path to discovery and a rigorous assessment of progress toward understanding the processes involved.
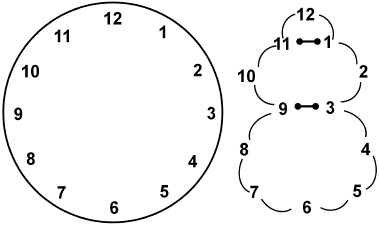
Figure 3.4
Basic DNA origami. One long (single-stranded) DNA circle is represented as a twelve-hour clock face on the left. The addition of two (single-stranded) staple oligos connect (via base-pairing each segment of the circle) 11 to 1 and 9 to 3.This results in a shape resembling a Dali-esque odalisque with a double-stranded collar and belt.
Between 1977 and 1981, based on the first folded RNA structure (tRNA,
Figure 3.1
), Ned Seeman and I invented ways to design and establish morphology from the basic base pairing rules of RNA and DNA (G with C, and A with U/T,
Figure 1.4
). We can now design and build atomically precise shapes swiftly and with generally high yield. We call these shapes DNA nanostructures (from Ned Seeman and William Shih), or DNA origami (from Paul Rothemund).
As a further example, 170 DNA staples (40-mers) bind to precise locations around a 7,000 base long single-stranded circle with half of each 40-mer staple binding one place and the other half binding somewhere else (designed by caDNAno, a computer-aided design process).
Shawn Douglas, Ido Bachelet, and I at the Wyss Institute for Biologically Inspired Engineering at Harvard have used this technique to construct nano robotsâessentially cages made of DNA that hold cancer-killing antibodies. These nano cages open up and release their cargo of antibodies only when they touch cancer cells. The rules for defining the shape of protein
structures are considerably harder than those for designing DNA structures (the awesome base pairs of
Figure 1.4
). The rules for designing cell and multicellular shapes are harder yet (unless, of course, we've seen them before).
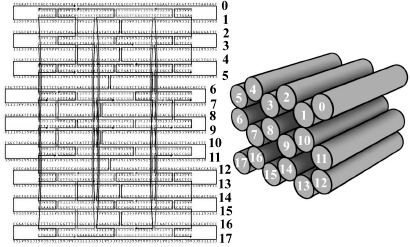
Figure 3.5
3-D DNA origami. One long DNA circle (represented as mostly horizontal lines plus far left and far right loops) is guided to fold into a specific 3-D log pile of eighteen double helices. The short, linear staples (mostly vertical lines plus internal loops) connect specific distant points on the circle.
Despite the three advantages listed above, multicellularity tends to come with a hard consequenceâyou give up your immortality. Most single-cell organism species seem to live indefinitely, its members still evolving, traceable as a continuous genetic lineage (especially in the highly conserved ribosomal RNA genes) back to the dawn of time. Selection in species with diverse cell types comes along with planned obsolescence and a circle of life progressionâfrom egg to chicken to egg. As single-cell organisms replicate by simple cell division, lessons learned (mutations and environmental responses) just prior to replication are often retained in the daughter cells. But with multicellular organisms, the larger they are, the more phases they have to pass through in going from egg to adult. In species that learn, a lifetime of
experience is lost at death. This loss can be partially compensated for by inheritance of cultural (non-DNA) artifacts, such as in the
Cyanistes
birds in the 1960s that taught each other how to open foil-topped British milk bottles.
In any case, the advantages and disadvantages of multicellularity may no longer apply to humans because of the huge impact of cultural inheritance and the prescient design of our technological culture. The advantages of multicellularity (reassortment of genes, division of labor and shapes) are now affected by markets and software. The downside of knowledge lost at death is offset by vast libraries and education, but a full education can take sixty years. So we'd like to either speed up education or slow down decay once a person is educated. The former might be made possible by making more rapid and accurate bridges between human knowledge and computer knowledge, either by the optimal use of existing senses or by means of multi-electrode neuronal input and output. Retarding the processes of decay, by contrast, might be made possible by discovering why some animals live long and vigorous lives while others die quickly (ranging from 3 days to 400 years; see
Chapter 9
). To study this phenomenon, we'd like to have two species that are closely related but have radically different life spans. As for example . . .