The Ghost in the Machine (26 page)
evolution of our own species. It is now generally recognised that the
human adult resembles more the embryo of an ape than an adult ape. In
both simian embryo and human adult, the ratio of the weight of the brain
to total body weight is disproportionately high. In both, the closing
of the sutures between the bones of the skull is retarded to permit
the brain to expand. The back-to-front axis through man's head -- i.e.,
the direction of his line of sight -- is at right angles to his spinal
column: a condition which, in apes and other mammals, is found only in the
embryonic, not in the adult stage. The same applies to the angle between
backbone and uro-genital canal -- which accounts for the singularity of
the human way of copulating face to face. Other embryonic -- or, to use
Bolk's term,
foetalised
-- features in adult man are: the absence of
brow-ridges; the scantness and late appearance of body hair; pallor of
the skin; retarded growth of the teeth, and a number of other features --
including 'the rosy lips of man which were probably evolved in the young
as an adaptation to prolonged suckling and have persisted in the adult,
possibly under the influence of sexual selection' (de Beer). [6]
wrote J.B.S. Haldane, 'it will probably involve still greater prolongation
of childhood and retardation of maturity. Some of the characters
distinguishing adult man will be lost.' [7] There is, incidentally, a
reverse of the medal which Aldous Huxley pointed out in one of his later,
despairing novels: artificial prolongation of the absolute lifespan of
man might provide an opportunity for features of the adult primate to
reappear in human oldsters: Methuselah would turn into a hairy ape.*
But this ghastly perspective does not concern us here.
* Huxley, After Many a Summer. Some physical characteristics in
the very old seem to indicate that the genes which could produce
such a transformation are still present in our gonads, but are
prevented from becoming active by the neotenic retardation of the
biological timeclock. The obvious conclusion is that prolongation
of the human lifespan is only desirable if it can be accompanied
by techniques which exert a parallel influence on the genetic clock.
retreat from specialised adult forms of bodily structure and behaviour,
to an earlier or more primitive, but also more plastic and less committed
stage -- followed by a sudden advance in a new direction. It is as
if the stream of life had momentarily reversed its course, flowing
uphill for a while, then opened up a new stream-bed. I shall try to
show that this
reculer pour mieux sauter
-- of drawing back to leap,
of undoing and re-doing -- is a favourite gambit in the grand strategy
of the evolutionary process; and that it also plays an important part
in the progress of science and art.
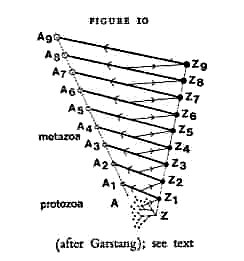
is meant to represent the progress of evolution by paedomorphosis. Z
to Z9 is the progression of zygotes (fertilised eggs) along the
evolutionary ladder; A to A9 represents the adult forms resulting from
each zygote. Thus the black line from Z4 to A4, for instance, represents
ontogeny, the transformation of egg into adult; the dotted line from A to
A9 represents phylogeny -- the evolution of higher forms. But note that
the thin lines of evolutionary progress do not lead directly from, say, A4
to A5 -- that would be gerontomorphosis, the evolutionary transformation
of an adult form. The line of progress branches off from the unfinished,
embryonic stage of A4. This represents a kind of evolutionary retreat
from the finished product, and a new departure towards the evolutionary
novelty Z5-A5. A4 could be the adult sea cucumber: then the branching-off
point on the line A4-Z4 would be its larva; or A8 could be the adult
primate ancestor of man, and the branching-off point its embryo --
which is so much more like the A9 -- ourselves.
the evolution of
ideas
. The emergence of biological novelties
and the creation of mental novelties are processes which show certain
analogies. It is of course a truism that in mental evolution, social
inheritance through tradition and written records replaces genetic
inheritance. But the analogy goes deeper: neither biological evolution
nor mental progress follows a continuous line from A6 to A7. Neither of
them is strictly cumulative in the sense of continuing to build where the
last generation has left off. Both proceed in the zigzag fashion indicated
in the diagram. The revolutions in the history of science are successful
escapes from blind alleys. The evolution of knowledge is continuous only
during those periods of consolidation and elaboration which follow a major
break-through. Sooner or later, however, consolidation leads to increasing
rigidity, orthodoxy, and so into the dead end of overspecialisation --
to the koala bear. Eventually there is a crisis and a new 'break-through'
out of the blind alley -- followed by another period of consolidation,
a new orthodoxy, and so the cycle starts again.
is not built on top of the previous edifice; it branches out from the
point where progress has gone wrong. The great revolutionary turns in
the evolution of ideas have a decidedly paedomorphic character. Each
zygote in the diagram would represent the seminal idea, the seed out of
which a new theory develops until it reaches its adult, fully matured
stage. One might call this the ontogeny of a theory. The history of
science is a series of such ontogenies. True novelties are not derived
directly from a previous adult theory, but from a new seminal idea --
not from the sedentary sea urchin but from its mobile larva. Only in
the quiet periods of consolidation do we find gerontomorphosis -- small
improvements added to a fully grown, established theory.
in evidence: Garstang's diagram could have been designed to show how
periods of cumulative progress within a given 'school' and technique
end inevitably in stagnation, mannerism or decadence, until the crisis
is resolved by a revolutionary shift in sensibility, emphasis, style.*
* See The Act of Creation, Book One, Chapters X and XXIII.
that it has a solid factual basis. Biological evolution is to a large
extent a history of escapes from the blind alleys of overspecialisation,
the evolution of ideas a series of escapes from the bondage of mental
habit; and the escape mechanism in both cases is based on the principle
of undoing and re-doing, the draw-back-to-leap pattern.
our starting point, the monkey at the typewriter. The monkey, according
to the orthodox doctrine, is supposed to proceed by hit and miss, just
as mental evolution, according to Behaviourist doctrine, is supposed
to proceed by trial and error. In both cases, progress is secured by
the stick-and-carrot method: the successful tries are rewarded by the
carrot of survival or of 'reinforcement'; the harmful ones are weeded
out by the stick of extinction, or by 'negative reinforcement'.
error are inherent in all progressive development. But there is a world
of difference between the random tries of the monkey at the typewriter,
and the various directive processes summarised in preceding chapters --
starting with the hierarchic controls and regulations built into the
genetic system, and culminating in the draw-back-to-leap pattern of
paedomorphosis. The orthodox view implies reeling off the available
responses in the animal's repertory, or on the Tibetan prayer-wheel of
mutations, until the correct one is hit upon by chance. The present view
also relies on trial and error -- each escape from a blind alley followed
by a new departure is just -- that but of a more complex, sophisticated
and purposive kind: a groping and exploring, retreating and advancing
towards higher levels of existence. 'Purpose', to quote H.J. Muller again,
'is not imported into Nature. . . . It is simply implicit in it.' [8]
known for some time, but their implications have mostly been ignored
by orthodox evolutionists. Yet if these isolated facts and theories
are worked into a synthesis, they make the problem of evolution
appear in a new light. There may be a monkey hammering away at the
typewriter, but that device is organised in such a way as to defeat
the monkey. Evolution is a process with a fixed code of rules, but
with adaptable strategies. The code is inherent in the conditions
of our planet; it restricts progress to a limited number of avenues;
while at the same time all living matter strives towards the optimal
utilisation of the offered possibilities. The combined action of these two
factors is manifested on each successive level: in the micro-hierarchy
of the gene-complex, the canalisation of embryonic development, and
its stabilisation by developmental homeostasis. Homologue organs --
evolutionary holons -- and similar animal forms arise from independent
origins and provide archetypal unity-in-variety. The initiative of
the animal, its curiosity and exploratory drive, act as pacemakers of
progress; a quasi-Lamarckian mechanism of inheritance may in rare cases
come to its aid; paedomorphosis offers an escape from blind alleys and a
new departure in a different direction; and lastly, Darwinian selection
operates within its limited scope.
trigger which releases the coordinated action of the system; and to
maintain that evolution is the product of blind chance means to confuse
the simple action of the trigger with the complex, purposive processes
which it sets off. Their purposiveness is manifested in different ways on
different levels of the hierarchy; on each level there is trial and error,
but on each level it takes a more sophisticated form. Some years ago,
two eminent experimental psychologists, Tolman and Krechevsky, created
a stir by proclaiming that the rat learns to run a maze by forming
hypotheses. [9] Soon it may be permissible to extend the metaphor and
to say that evolution progresses by making and discarding hypotheses,
in the process of spelling out a roughed-in idea.
We are all in the gutter,
but some of us are looking at the stars.
Oscar Wilde
ingenious improvisations, according to the challenge they face.* Other
things being equal, a monotonous environment leads to the mechanisation of
habits, to stereotyped routines which, repeated under the same unvarying
conditions, follow the same rigid, unvarying course. The pedant who has
become a slave of his habits thinks and acts like an automaton running
on fixed tracks; his biological equivalent is the over-specialised animal
the koala bear clinging to his eucalyptus tree.
* See Chapter Eight.
which can only be met by flexible behaviour, variable strategies,
alertness for exploiting favourable opportunities. The biological parallel
is provided by the evolutionary strategies discussed in previous chapters.
no longer be met by the organism's customary skills. In such a major
crisis -- and both biological evolution and human history are punctuated
by such crises -- one of two possibilities may occur. The first is
degenerative
-- leading to stagnation, biological senescence,
or sudden extinction as the case may be. In the course of evolution this
happened over and again; to each surviving species there are a hundred
which failed to pass the test.
Part Three
of this book discusses the possibility that our own species is facing
a crisis unique in its history, and that it is in imminent danger of
failing the test.
regenerative
Other books
Quake by Jacob Chance
Monroe, Marla - Desire for Two (Siren Publishing Ménage Amour) by Marla Monroe
How to Break a Terrorist by Matthew Alexander
Twelve Desperate Miles by Tim Brady
Nothing In Her Way by Charles Williams
The Best American Mystery Stories, Volume 17 by Lisa Scottoline
Binder - 02 by David Vinjamuri
A Path to Coldness of Heart by Glen Cook
The Robber Bridegroom by Welty,Eudora.
Finding Hope by Broas, K