Power, Sex, Suicide: Mitochondria and the Meaning of Life (15 page)
Read Power, Sex, Suicide: Mitochondria and the Meaning of Life Online
Authors: Nick Lane
Tags: #Science, #General
Keilin and Warburg maintained a lively correspondence throughout the 1920s and 1930s, disagreeing on many particulars. Ironically, Keilin’s concept of a respiratory chain was given more credence by Warburg himself, who discovered additional non-protein components of the chain during the 1930s, which are now referred to as coenzymes. For his new discoveries Warburg was offered a second Nobel Prize in 1944, but, being Jewish, he was refused permission to receive it by Hitler (who nonetheless allowed himself to be swayed by Warburg’s international prestige, and did not have him imprisoned, or worse). Sadly, Keilin’s own profound insights into the structure and function of the respiratory chain were never honoured with a Nobel Prize, surely an oversight on the part of the Nobel committee.
The overall picture then emerging was this. Glucose is broken down into smaller fragments, which are fed into an asset-stripping merry-go-round of linked reactions, known as the Krebs Cycle.
2
These reactions strip out the carbon and oxygen atoms, and discharge them as carbon dioxide waste. The hydrogen atoms bind to Warburg’s coenzymes and enter the respiratory chain. There, the hydrogen atoms are split into their constituent electrons and protons, the further passage of which differs. We’ll look into what happens to the protons later on; for now, we’ll concentrate on the electrons. These are passed down the full length of the chain by the string of electron carriers. Each of the carriers is successively reduced (gaining electrons) and then oxidized (losing electrons) by the next link in the chain. This means that the respiratory chain forms a succession of linked redox reactions, and so behaves like a tiny electrical wire. Electrons are transferred down the wire from carrier to carrier at a rate of about 1 electron every 5 to 20 milliseconds. Each of the redox reactions is exergonic—in other words, each releases energy that can be used for work. In the final step, the electrons pass from cytochrome c to oxygen, where they are reunited with protons to form water. This last reaction takes place in Warburg’s respiratory ferment, which had been re-named
cytochrome oxidase
by Keilin, because it uses oxygen to oxidize cytochrome c. Keilin’s term is still in use today.
Today we know that the respiratory chain is organized into four gigantic molecular complexes, embedded in the inner membrane of the mitochondria (
Figure 5
). Each complex is millions of times the size of a carbon atom, but even so they are barely visible down the electron microscope. The individual complexes are composed of numerous proteins, coenzymes, and cytochromes, including those discovered by Keilin and Warburg. Curiously, mitochondrial genes encode some of the proteins, while nuclear genes encode others, so the complexes are an amalgam encoded by two separate genomes. There are tens of thousands of complete respiratory chains embedded in the inner membrane of a single mitochondrion. It seems that these chains are physically separated from each other, and indeed even the complexes within individual chains seem to be physically independent.
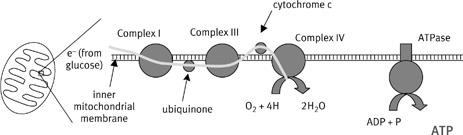
5
Simplified representation of the respiratory chain, showing complexes I, III, and IV, and the ATPase. Complex II is not shown here, as electrons (e
–
) enter the chain at either complex I or complex II, and are passed on from either of these complexes to complex III by the carrier ubiquinone (also known as Coenzyme Q, sold in supermarkets as a health food supplement, though with questionable efficacy). The passage of electrons down the chain is illustrated by the curvy line. Cytochrome c carries electrons from complex III to complex IV (cytochrome oxidase) where they react with protons and oxygen to form water. Notice that all of the complexes are embedded separately in the membrane. Whereas ubiquinone and cytochrome c shuttle electrons between the complexes, the nature of the intermediate that connected electron flow down the respiratory chain with ATP synthesis in the ATPase was a mystery that confounded the field for an entire generation.
Although Keilin’s early concept of the respiratory chain was correct in its essentials, perhaps the most important question remained unanswered—how was energy conserved, rather than being dissipated on the spot? Energy is released by the passage of electrons down the respiratory chains to oxygen, but is consumed elsewhere in the cell, usually outside the mitochondria, at a later time. There had to be some kind of intermediate, presumably a molecule of some sort, that could conserve the energy released in respiration, then transfer it to other compartments of the cell, and couple it to some kind of work. Whatever this intermediate was, it had to be sufficiently adaptable to be used for the various different types of work carried out by the cell, and it had to be stable enough to remain intact until needed (as even moving around the short distances of the
cell takes some time). In other words, it had to be the molecular equivalent of a common currency, or coins that could be exchanged for services rendered. The respiratory chain is then the Mint, where the new currency is produced. So what could this currency be?
The first glimmer of an answer came from studies of fermentation. The ageold importance of fermentation in wine-making and brewing belied how little was known about the process. The beginnings of a chemical understanding came from Lavoisier, once again, who measured the weight of all the products, and declared fermentation to be no more than a chemical splitting of sugar to give alcohol and carbon dioxide. He was quite right, of course, but in a sense he missed the point, for Lavoisier thought of fermentation as just a chemical process with no inherent function. For Lavoisier, yeast was merely sediment that happened to catalyse the chemical breakdown of sugar.
By the nineteenth century, the students of fermentation split into two camps—those who thought that fermentation was a living process with a function (mostly the vitalists, who believed in a special vital force, irreducible to ‘mere’ chemistry), and those who considered fermentation to be purely a chemical process (mostly the chemists themselves). The century-long feud appeared to be settled by Louis Pasteur, a vitalist, who demonstrated that yeast was composed of living cells, and that fermentation was carried out by these cells in the absence of oxygen. Indeed Pasteur famously described fermentation as ‘life without oxygen’. As a vitalist, Pasteur was convinced that fermentation must have a purpose, which is to say, a function that was beneficial in some way for yeast, but even he admitted to being ‘completely in the dark’ about what this purpose might have been.
Only two years after Pasteur’s death in 1895, the belief that living yeast was necessary for fermentation was overturned by Eduard Buchner, who received the Nobel Prize for his work in 1907. Buchner used German brewers’ yeast rather than Pasteur’s French vintners’ yeast. Clearly the German yeast was more robust, for unlike Pasteur, Buchner did succeed in grinding it up with sand in a mortar to form a paste, then squeezing juice from the paste using a hydraulic press. If sugar was added to this ‘pressed yeast juice’, and the mixture incubated, fermentation began within a few minutes. The mixture evanesced alcohol and carbon dioxide in the same proportions as live yeast, albeit in lesser volume. Buchner proposed that fermentation was carried out by biological catalysts that he named
enzymes
(from the Greek
en zyme
, meaning
in yeast
). He concluded that living cells are chemical factories, in which enzymes manufacture the various products. For the first time, Buchner had shown that these chemical factories could be reconstituted even after the demise of the cells themselves, so long as the conditions were suitable. This discovery heralded the end of vitalism, and a new sense that all living processes could ultimately
be explained by similarly reductionist principles—the dominant theme of twentieth century biomolecular sciences. But Buchner’s legacy also reduced living cells to little more than a bag full of enzymes, which to this day dulls our perception of the importance of membranes in biology, as we shall see.
Using Buchner’s yeast juices, Sir Arthur Harden in England and Hans von Euler in Germany (and others) gradually pieced together the succession of steps in fermentation during the first decades of the twentieth century. They unravelled about a dozen steps in all, each catalysed by its own enzyme. The steps are linked together like a factory production line, in which the product of one reaction is the starting point for the next. For their work, Harden and von Euler shared the Nobel Prize in 1929. But the biggest surprise came in 1924, when yet another Nobel laureate, Otto Meyerhof, showed that almost exactly the same process takes place in muscle cells. In muscles, admittedly, the final product was lactic acid, which produces cramps rather than the enjoyable inebriation of alcohol, but Meyerhof showed that almost all the twelve factory production-line steps are the same. This was a striking demonstration of the fundamental unity of life, implying that even simple yeasts are related by descent to human beings, as postulated by Darwin.
By the end of the 1920s it was becoming clear that cells use fermentation to generate energy. Fermentation acts as a backup power supply (indeed the only power supply in some cells), which is usually switched on when the main energy generator, oxygen respiration, fails. Thus, fermentation and respiration came to be seen as parallel processes, both of which served to provide energy for cells, one in the absence of oxygen, the other in its presence. But the larger question remained: how are the individual steps coupled to the conservation of energy for use in other parts of the cell, at other times? Did fermentation, like respiration, generate some kind of energy currency?
The answer came in 1929 with the discovery of ATP by Karl Lohman in Heidelberg. Lohman showed that fermentation is linked to the synthesis of ATP (adenosine triphosphate), which can be stored in the cell for use over a period of hours. ATP is composed of adenosine bound to three phosphate groups linked end to end in a chain, a somewhat precarious arrangement. Splitting off the terminal phosphate group from ATP releases a large amount of energy that can be used to power work—indeed, is
required
to power much biological work. In the 1930s the Russian biochemist Vladimir Engelhardt showed that ATP is necessary for muscle contraction—muscles tense in a state of rigor, as in rigor mortis, when deprived of ATP. Muscle fibres split ATP to liberate the energy that they need to contract and relax again, leaving adenosine diphosphate (ADP) and phosphate (P):
ATP → ADP + P + energy.
Because the cell has a limited supply of ATP, fresh supplies must be regenerated continually from ADP and phosphate—and to do so, of course, requires an input of energy, as can be seen if the equation above is followed backwards. This is the function of fermentation: to provide the energy that is needed to regenerate ATP. Fermenting one molecule of glucose regenerates two molecules of ATP.
Engelhardt immediately squared up to the next question. ATP is
needed
for muscle contraction, but is only produced by fermentation when oxygen levels are low. If the muscles are to contract in the
presence
of oxygen, then presumably some other process must generate the ATP needed; this, said Engelhardt, must be the function of oxygen respiration. In other words, oxygen respiration also serves to generate ATP. Engelhardt set about trying to prove his assertion. The difficulty faced by researchers at the time was technical: muscles are not easy to grind up to use for studies of respiration—they are damaged and leak. Engelhardt resorted to an unusual experimental model, which could be manipulated rather more easily—the red blood cells of birds. Using them, he showed that respiration really does generate ATP, and in far greater quantities than fermentation. Soon afterwards, the Spaniard Severo Ochoa showed that as many as 38 molecules of ATP could be generated by respiration from a single molecule of glucose, a finding for which he too received the Nobel Prize, in 1959. This means that oxygen respiration can produce 19 times more ATP per molecule of glucose than does fermentation. The total production is astonishing. In an average person, ATP is produced at a rate of 9 × 10
20
molecules per second, which equates to a turnover rate (the rate at which it is produced and consumed) of about
65 kg
every day.
Few people accepted the universal significance of ATP at first, but work by Fritz Lipmann and Herman Kalckar in Copenhagen in the 1930s confirmed it, and by 1941 (now in the US), they proclaimed ATP to be ‘the universal energy currency’ of life. In the 1940s this must have been an audacious claim, the kind that can easily backfire and cost the advocates their careers. Yet astonishingly, given the flamboyance and variety of life, it is basically true. ATP has been found in every type of cell ever studied, whether plant, animal, fungal, or bacterial. In the 1940s, ATP was known to be the product of both fermentation and respiration, and by the 1950s, photosynthesis was added to the list—it, too, generates ATP, in this case by trapping the energy of sunlight. So the three great energy highways of life, respiration, fermentation, and photosynthesis, all generate ATP, another profound example of the fundamental unity of life.