Power, Sex, Suicide: Mitochondria and the Meaning of Life (21 page)
Read Power, Sex, Suicide: Mitochondria and the Meaning of Life Online
Authors: Nick Lane
Tags: #Science, #General
Ultimately, biological fitness is written in the sequence of the genes, for they alone are passed on to the next generation (well, almost alone: mitochondria are, too). Over evolutionary time, alterations in the genetic sequence, subjected to round after round of natural selection, build tiny refinement upon tiny refinement, until finally erecting the dizzying cathedral of biological complexity. Although Darwin knew nothing of genes, the genetic code at once suggests a mechanism for producing random variation in a population: mutations in the sequence of ‘letters’ in DNA can change the sequence of amino acids in proteins, which might have a positive, or a negative, or a neutral, effect on their function. Copying errors alone generate such variation. Each generation produces perhaps several hundred small changes in the DNA sequence (out of several billion letters), which may or may not affect fitness. Such small changes undoubtedly occur, and generate some of the raw material for the slow evolutionary change anticipated by Darwin. The gradual divergence in the sequence of genes of different species, over hundreds of millions of years, shows this process in action.
But small mutations are not the only way to bring about change in the genome (the complete library of genes in one organism), and the more we learn about genomics (the study of genomes), the less important small mutations seem to be. At the least, greater complexity demands more genes—the small bacterial genome could hardly code for a whole human being, still less the
myriad genetic differences between individuals. Surveying species, there is a general correlation between the degree of complexity and the number of genes, if not the total DNA content. So where do all these extra genes come from? The answer is duplications of existing genes, or whole genomes, or from the union of two or more different genomes, or from the spread of repetitive DNA sequences—apparently ‘selfish’ replicators, which copy themselves throughout the genome, but may later be co-opted to serve some useful function (useful, that is, to the organism as a whole).
None of these processes is strictly Darwinian, in the sense of gradual, small refinements to an existing genome. Rather, they are large-scale, dramatic changes in the total DNA content—giant leaps across genetic space, transforming existing gene sequences at a single stroke—even if they generate the raw material for new genes, rather than the new genes themselves. Excepting these leaps across genetic space, the process is otherwise Darwinian. Changes to the genome are brought about in an essentially random manner, and then subjected to rounds of natural selection. Small changes hone the sequence of new genes to new tasks. So long as the big jumps in DNA content do not generate an unworkable monster, they can be tolerated. If there is no benefit in having twice as much DNA, then we can be sure that natural selection will jettison it again—but if complex organisms need a lot of genes, then the elimination of superfluous DNA surely puts a ceiling on the maximum possible complexity, for it eliminates the raw material needed to form new genes.
This brings us back to the ramp of complexity. We have seen that there is a big discontinuity between bacteria and eukaryotes. It is remarkable that bacteria are
still
bacteria: while enormously varied and sophisticated in biochemical terms, they have resolutely failed to generate real
morphological
complexity in four billion years of evolution. In their size, shape, and appearance, they can hardly be said to have evolved in any direction at all. In contrast, in half the time open to bacteria, the eukaryotes unquestionably ascended a ramp of complexity—they developed elaborate internal membrane systems, specialized organelles, complex cell cycles (rather than simple cell division), sex, huge genomes, phagocytosis, predatory behaviour, multicellularity, differentiation, large size, and finally spectacular feats of mechanical engineering: flight, sight, hearing, echolocation, brains, sentience. Insofar as this progression happened over time, it can reasonably be plotted out as a ramp of ascending complexity. So we are faced with bacteria, which have nearly unlimited biochemical diversity but no drive towards complexity, and eukaryotes, which have little biochemical diversity, but a marvellous flowering in the realm of bodily design.
When confronted with the divide between bacteria and eukaryotes, the Darwinian might reply: ‘Ah, but the bacteria
did
generate complexity—they
gave rise to the more complex eukaryotes, which in turn gave rise to many organisms of inordinately greater complexity.’ This is true, but only in a sense, and here is the rub. The mitochondria, I shall argue, could
only
be derived by endosymbiosis—a union of two genomes in the same cell, or a giant leap across genetic space—and without mitochondria, the complex eukaryotic cell simply could
not
evolve. This viewpoint stems from the idea that the eukaryotic cell itself was forged in the merger that gave rise to mitochondria, and that the possession of mitochondria is, or was in the past, a
sine qua non
of the eukaryotic condition. This picture differs from the mainstream view of the eukaryotic cell, so let’s remind ourselves quickly why it matters.
In
Part 1
, we examined the origin of the eukaryotic cell, as surmised by Tom Cavalier-Smith, which best represents the mainstream view. To recapitulate, a prokaryotic cell (without a nucleus) lost its cell wall, perhaps through the action of an antibiotic produced by other bacteria, but survived the loss, as it already had an internal protein skeleton (cytoskeleton). The loss of the cell wall had profound consequences for the cell in terms of its lifestyle and manner of reproduction. It developed a nucleus and a complicated life cycle. Using its cytoskeleton to move around and change shape like an amoeba, it developed a new, predatory lifestyle, engulfing large particles of food such as whole bacteria by phagocytosis. In short, the first eukaryotic cell evolved its nucleus and its eukaryotic lifestyle by standard Darwinian evolution. At a relatively late stage, one such eukaryotic cell happened to engulf a purple bacterium, perhaps a parasite like
Rickettsia
. The internalized bacteria survived and eventually transmuted, by standard Darwinian evolution, into mitochondria.
Notice two things about this line of reasoning: first, it exhibits what we might call a Darwinian bias, in that it limits the importance attributed to the union of two dissimilar genomes, a basically non-Darwinian mode of evolution; and second, it limits the importance of mitochondria in this process. Mitochondria are incorporated into a fully functional eukaryotic cell, and are readily lost again in many primitive lines such as
Giardia
. Mitochondria, in this view, are an efficient means of generating energy, but no more nor less than that. The new cell simply had a Porsche engine fitted, in place of its old-fashioned milk-cart motor. I think this view gives little real insight into why all complex cells possess mitochondria, or conversely, why mitochondria are
needed
for the evolution of complexity.
Now consider the hydrogen hypothesis of Bill Martin and Miklos Müller, which we also discussed in
Part 1
. According to this radical hypothesis, a mutual chemical dependency between two very different prokaryotic cells led to a close relationship between the two. Eventually one cell physically engulfed the other, combining two genomes within a single cell: a giant leap across genetic space to create a ‘hopeful monster’. This genetic leap, in turn, set up a series of
Darwinian selection pressures on the new entity, leading to a transfer of genes from the guest to the host. The critical point of the hydrogen hypothesis is that there
never was
a primitive eukaryote, one that supposedly possessed a nucleus and had a predatory lifestyle, but did not have any mitochondria. Rather, the first eukaryote was born of the union between two prokaryotes, a fundamentally non-Darwinian process—there was no halfway house.
Just look at
Figure 9
, a tree of life drawn in 1905 by the Russian biologist Konstantine Merezhkovskii, to see what an uncomfortable reversal of the standard branching tree of life this creates. There has been plenty of controversy over trees of life in the past, notably from Stephen Jay Gould, who claimed that the Cambrian explosion inverted the usual tree. The Cambrian explosion refers to the great, and geologically sudden, proliferation of life around 560 million years ago. Later on, most of the major branches were ruthlessly pollarded, as whole phyla fell extinct. Daniel Dennett, in
Darwin’s Dangerous Idea
, lambasts Gould’s apparently radical evolutionary trees for being the same as any other evolutionary tree, except with distorted axes—a low-lying scrub bush, throwing up a few scraggly shoots, rather than a lofty tree of life. But there is no danger of this in Merezhkovskii’s case. His evolutionary tree is a genuinely upside down variety. Here, the branches fuse, rather than bifurcate, to generate a new domain of life.
I’m not trying to cry revolution. There is nothing exceptional about these arguments, and symbiosis is part of the standard evolutionary canon, even if it is played down as a mechanism of generating novelty. For example, the late, great John Maynard Smith and Eörs Szathmáry, in their stimulating book
The Origins of Life
, argue that biological symbiosis is analogous to a motorbike, which is a symbiosis between a bicycle and the internal combustion engine. Even if we view this symbiosis as an advance, they say, with rather crusty humour, someone still had to invent the bicycle and the internal combustion engine first. Likewise in life, natural selection must invent the parts first, and symbiosis just makes creative use of the available parts. Thus symbiosis is best explained in Darwinian terms.
All this is true, but it obscures the fact that some of the most profound evolutionary novelties are made possible
only
by symbiosis. Presumably, if we follow Maynard Smith and Szathmáry, if a bicycle and an internal combustion engine can evolve independently by natural selection, then so too, in principle, could the motorcycle. No doubt it’s faster to evolve a motorcycle by shuffling existing components, but there is no fundamental reason why it should not have evolved anyway, given enough time, in the absence of symbiosis. In the case of the eukaryotic cell, I disagree. Left to themselves, I will argue, bacteria could not evolve into eukaryotes by natural selection alone: symbiosis was
needed
to bridge the gulf between bacteria and eukaryotes, and in particular a mitochondrial merger was
necessary
to sow the seeds of complexity. Without mitochondria, complex life is simply not possible, and without symbiosis, mitochondria are not possible—without the mitochondrial merger we would be left with bacteria and nothing but. Regardless of whether we consider symbiosis Darwinian or not, an understanding of why symbiotic mitochondria are necessary is paramount to an understanding of our own past, and our place in the universe.
1
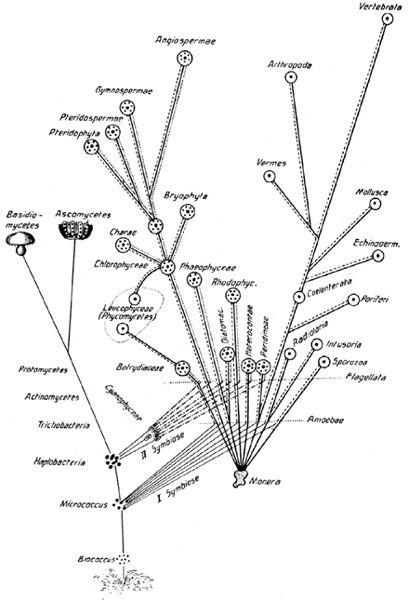
9
Merezhkovskii’s inverted tree of life, showing fusion of branches. The standard ‘Darwinian’ tree of life is strictly bifurcating: branches branch but do not fuse. The origin of the eukaryotic cell was endosymbiotic. On the tree of life, this is represented as bifurcation backwards: branches fuse together, inverting part of the tree of life.
In
Part 3
, we will see why there is such a yawning chasm between the prokaryotes and the eukaryotes, and why this deep divide can only be bridged by symbiosis—it is next to impossible, given the mechanism of chemiosmotic energy production (discussed in
Part 2
) for eukaryotes to evolve by natural selection from prokaryotes. This is why bacteria are still bacteria, and why it is unlikely that life as we know it, based on cells, carbon chemistry, and chemiosmosis, will progress beyond the bacterial level of complexity anywhere else in the universe. In
Part 3
, we’ll see why mitochondria seeded complexity in the eukaryotes, placing them at the beginning of the ramp of ascending complexity; and in
Part 4
, we’ll see why mitochondria impelled the eukaryotes onwards up the ramp.
Why Bacteria are Simple
The great French molecular biologist François Jacob once remarked that the dream of every cell is to become two cells. In our own bodies, this dream is held very carefully in check; otherwise the result would be cancer. But Jacob was trained as a microbiologist, and for bacteria, one cell becoming two is more than a dream. Bacteria replicate at colossal speed. When well fed,
E. coli
bacteria divide once every 20 minutes, or 72 times a day. A single
E. coli
bacterium weighs about a trillionth of a gram (10
–12
g). Seventy-two cell divisions in a day corresponds to an amplification of 2
72
(= 10
72 × log2
= 10
21.6
), which is an increase in weight from 10
–12
grams to 4000 metric tons. In two days, the mass of exponentially doubling
E. coli
would be 2664 times larger than the mass of the Earth (which weighs 5.977 × 10
21
metric tons)!