In The Blink Of An Eye (38 page)
Read In The Blink Of An Eye Online
Authors: Andrew Parker
Due to the short length of their stalks, the eyes of the Burgess arthropod
Sanctacaris
are greatly restricted in that they can only be directed forwards, suggesting a predatory lifestyle. And then another Burgess arthropod,
Yohoia
, has eyes fixed in position with bulbous, enlarged regions directed forwards, again further suggesting that predators were in existence 515 million years ago. There are other signs of predation in the Burgess Shale fossils, which will form the subject of the next part of this chapter, but first we could consider the Cambrian trilobites, which often show details of the individual facets of their compound eyes.
Sanctacaris
are greatly restricted in that they can only be directed forwards, suggesting a predatory lifestyle. And then another Burgess arthropod,
Yohoia
, has eyes fixed in position with bulbous, enlarged regions directed forwards, again further suggesting that predators were in existence 515 million years ago. There are other signs of predation in the Burgess Shale fossils, which will form the subject of the next part of this chapter, but first we could consider the Cambrian trilobites, which often show details of the individual facets of their compound eyes.
Most trilobite eyes, particularly the holochroal eyes that were the first to appear on Earth, have larger facets at their centres than at their edges. The eyes of early trilobites were positioned on the sides of the head, but were curved to scan the complete horizon around them. So they saw with greater precision towards their sides, at right angles to the forward direction of the trilobite. These characters appear contra-dictory
when compared with most living animals - eyes on the sides of the head suggest prey, while larger facets in the centre of the eye suggest predator. But there are animals in the sea today that do have eyes with similar directional properties to the trilobites - the fishes.
when compared with most living animals - eyes on the sides of the head suggest prey, while larger facets in the centre of the eye suggest predator. But there are animals in the sea today that do have eyes with similar directional properties to the trilobites - the fishes.
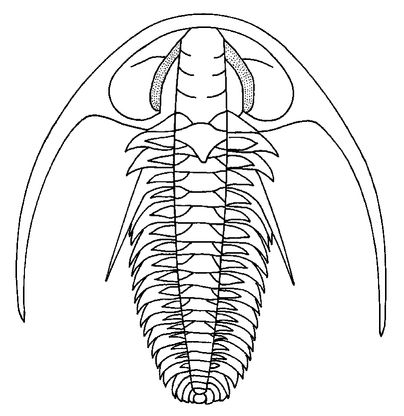
Figure 8.2
The early Cambrian trilobite
Fallotaspis typica
showing eyes (shaded) positioned at the side of the head, although its sight is directed slightly forward.
The early Cambrian trilobite
Fallotaspis typica
showing eyes (shaded) positioned at the side of the head, although its sight is directed slightly forward.
Fishes have eyes positioned at the sides of their head yet do not see equally in all directions. But fishes have camera-type and not compound eyes, so how can we infer such information when there is only one lens? The answer lies with the retina, and the distribution of the light detection cells within it.
If the eyeball of a fish is cut along its âequator', and the lower hemisphere is cut along its âlines of longitude', it can be laid flat. A globe is viewed on the flat pages of an atlas in this manner. The lower hemisphere of the eyeball is the retina, the region of light detection cells
where the image of the eye is formed. Objects to the side of the eye are imaged at the edges of the retina, while objects positioned along the central axis of the eye are imaged in the centre of the retina. The retina can be examined under microscopes to map the light detection cells. The results are always the same - the greatest concentration of light detection cells lies around the centre of the retina. Fishes see best along the central axis of their eyes, or out from the sides of the head.
where the image of the eye is formed. Objects to the side of the eye are imaged at the edges of the retina, while objects positioned along the central axis of the eye are imaged in the centre of the retina. The retina can be examined under microscopes to map the light detection cells. The results are always the same - the greatest concentration of light detection cells lies around the centre of the retina. Fishes see best along the central axis of their eyes, or out from the sides of the head.
Fishes can move their eyeballs within their sockets to some extent, but in comparison the region of high visual sensitivity in trilobites was larger than in fish, and so eye movements were probably not so necessary for trilobites to precisely track an object in the water. Considering that life supposedly functioned in the past as it does today, maybe trilobites were the fishes of the Cambrian.
These generalisations are obviously very broad. And fishes of open water can be predators, scavengers or herbivores, not to mention the fact that most are also potential prey. So this line of enquiry is, unfortunately, approaching the end of its usefulness, although it will be considered further in the following chapter. The ambiguity of eye position and architecture in relation to position in the food web means that we must look elsewhere for signs of predation in the Cambrian. And the most obvious place to look will turn out to be the best one - the whole fossilised bodies of the Cambrian animals themselves.
Swords, shields and scarsPost-Cambrian - the potential
So far I have been looking for secondary signs of predation. But what about primary signs - the knife-like tools and bite marks themselves? Rather than pursuing gun sights or the criminal mind, maybe we should be scouring the fossil record for the murder weapons and victims? And what of the shields used to protect from those weapons? It is worth searching for this evidence.
One of the most interesting museum specimens must be the âDeath of a goanna', the centrepiece of the Queensland Museum in Australia. This metre-long goanna, or monitor lizard, is preserved in its death
pose - mouth wide open and stuffed with an echidna. The goanna (half-wittedly) attempted to swallow the foot-long marsupial, complete with its outer coat of long spines. The spines punctured the mouth of the goanna in all directions, and the animals locked together and died in stalemate (see Plate 25).
pose - mouth wide open and stuffed with an echidna. The goanna (half-wittedly) attempted to swallow the foot-long marsupial, complete with its outer coat of long spines. The spines punctured the mouth of the goanna in all directions, and the animals locked together and died in stalemate (see Plate 25).
Chicago's Field Museum of Natural History displays a comparable specimen. Here the rear half of a herring-like prey emerges from the mouth of a perch-like predator. The herring proved to be more than a mouthful as both fish died in this irreversible position. But in this case the animals inhabited a lake 50 million years ago. They are preserved as fossils in a huge slab of limestone, recovered 2,500 metres above sea level in Wyoming.
It is rare to find ancient feeding preserved in action in the fossil record, but there are primary signs of both predators and prey locked within numerous fossils. Dinosaurs provide the obvious examples. The dentition of
T. rex
can mean only one thing - it ate meat. But did it kill or poach its meat - was it a predator or scavenger? The speed at which
T. rex
ran, as calculated from footprints, suggests it was capable of catching living prey. But this question remains a little contentious.
T. rex
can mean only one thing - it ate meat. But did it kill or poach its meat - was it a predator or scavenger? The speed at which
T. rex
ran, as calculated from footprints, suggests it was capable of catching living prey. But this question remains a little contentious.
In South Dakota, an amateur fossil hunter unearthed part of a bone from a
Hyracodon
, a thirty-million-year-old pig. Many other bones had been recovered from this extinct species, but there was something unusual about this one. Something was not quite right.
Hyracodon
, a thirty-million-year-old pig. Many other bones had been recovered from this extinct species, but there was something unusual about this one. Something was not quite right.
The bone was about the size of a golf ball. It was quite unexciting, except for some marks - clear, neat indentations up to a centimetre deep in places. Then the fossil hunter found a jaw from
Hoplophoneus
, a cat that lived in the same region as
Hyracodon
. The cat's dentition precisely matched the marks in the pig bone. It seemed conclusive that the cat had eaten the pig thirty million years ago. But when the ancient cat dined, was the pig already dead, or did the cat kill it? We will never know the answer. More telling, however, are the puncture marks in ammonoids.
Hoplophoneus
, a cat that lived in the same region as
Hyracodon
. The cat's dentition precisely matched the marks in the pig bone. It seemed conclusive that the cat had eaten the pig thirty million years ago. But when the ancient cat dined, was the pig already dead, or did the cat kill it? We will never know the answer. More telling, however, are the puncture marks in ammonoids.
The now extinct ammonoids, as featured in Chapters 2 and 6, lived within a hard, spiral shell, allowing their tentacles to protrude into the water. Ammonoids probably hunted in the manner of squids and cuttlefishes today. Here tentacles grip prey with their suckers, while their
beak-like mouthparts and file-like inner teeth perform the cutting and grinding. But we know something else about feeding involving ammonoids - this time as prey.
beak-like mouthparts and file-like inner teeth perform the cutting and grinding. But we know something else about feeding involving ammonoids - this time as prey.
During their reign, ammonoids swam successfully through ancient seas. Occasionally, however, they would be seen falling through the water, plunging to the ocean floor. These ammonoids were dead, or dying . . . but isn't this theory contradictory? When ammonoids died, the gases released from their decaying bodies inflated their shells. The buoyant shells then floated to the surface and were washed ashore, where they were laid to rest in the shallow ammonoid graveyard. Yet some sinking ammonoids were heading for a deep-water grave - but why?
Sometimes ammonoid shells
are
recovered from deep-water localities. Sometimes they
did
fall to the sea floor directly below their natural waters. The fossils in this case, however, are different from those of shallow-water graves. The shells found in the unnatural, shallow-water sites are intact. The shells recovered from their owners' original localities bear puncture marks.
are
recovered from deep-water localities. Sometimes they
did
fall to the sea floor directly below their natural waters. The fossils in this case, however, are different from those of shallow-water graves. The shells found in the unnatural, shallow-water sites are intact. The shells recovered from their owners' original localities bear puncture marks.
The puncture marks are roughly circular and the sizes of various coins. Cracks often radiate from these marks. Some shells have puncture marks that are randomly arranged, while others have marks arranged in patterns. There are two theories for the cause of the randomly arranged marks. The first is that limpets caused them.
Limpets are snails with hat-shaped shells. They graze on rocks or other hard surfaces. After grazing they often return to the same resting place, eventually forming a shallow, round depression. One idea is that the shells of ammonoids were suitably hard surfaces which ancient limpets could make their resting places. In this case, cracks radiating from the holes would be artefacts of deep burial and, consequently, high pressure. The alternative theory, however, is more dramatic, and certainly explains the regular patterns of puncture marks formed.
Mosasaurs were large, marine reptiles that lived alongside ammonoids. Their crocodile-like dentition suggests they were predators that patrolled the open water of ancient seas. But there is something else that can be inferred from their complete dentition - that they preyed upon ammonoids.
Jaws of mosasaurs can be found which explain the patterns of marks in ammonoid shells. When the shells are placed between the jaws of a certain size, the teeth fit precisely into the marks. The size and position of the mosasaur teeth within their jaws are a perfect match with the puncture marks of ammonoids. Case closed. Now we can re-reconstruct those ancient seas with ammonoids swimming, but this time with mosasaurs snapping at them.
Whether or not the randomly arranged puncture marks were made by limpets or mosasaurs, which may have taken several bites, they do explain the deep-water burial of these ammonoids. When punctured, the ammonoid shell would begin to fill with water, although the living part of the ammonoid would remain alive. As water infiltrated the otherwise gas-filled chambers of the shell, the ammonoid would become less buoyant and start to sink. Lying helpless on the sea floor or incapable of movement on its way there, the ammonoid would be susceptible to further, fatal attacks by the mosasaur. And the shell would remain below the scene of the crime, to be buried in a deep-water locality and not with those ammonoids that died a natural death, in their shallow-water graveyards. But their problems all began when their probable camouflage cover was blown - mosasaurs were visual hunters.
The 20,380-year-old Siberian mammoth first mentioned in Chapter 2 was found alone in frozen ground and French scientists have been investigating the cause of death of this specimen in the hope of explaining the extinction of mammoths in general. But maybe a single specimen will never provide the answer to this dilemma. Bones from
many
mammoths, however, were uncovered from a site in England, and it is the number of individuals that suggests the mammoth was victim of a successful hunting strategy. That, and the marks of predation.
many
mammoths, however, were uncovered from a site in England, and it is the number of individuals that suggests the mammoth was victim of a successful hunting strategy. That, and the marks of predation.
In a burial site for ancient Britons who lived up to 50,000 years ago (ending with, apparently, King Arthur), the bones of other animals are to be found. At Wookey Hole near Glastonbury in England there is an extensive system of caves. The entrance to these caves lies at the foot of a 50-metre, vertical cliff. At the foot of this cliff there is also a small recess, protected from the often harsh climate. Within this recess, two sets of bones have been found - those of predators and those of plant
eaters, or prey. The ancient predators of Wookey Hole were hyenas, and the prey were mainly mammoths. The hyena teeth fit precisely into marks in the mammoth bones, evidence that hyenas once preyed upon mammoths. But how could hyenas kill such a huge animal as a mammoth, and how did they lure the mammoths into their den, the recess in the cliff?
eaters, or prey. The ancient predators of Wookey Hole were hyenas, and the prey were mainly mammoths. The hyena teeth fit precisely into marks in the mammoth bones, evidence that hyenas once preyed upon mammoths. But how could hyenas kill such a huge animal as a mammoth, and how did they lure the mammoths into their den, the recess in the cliff?
Mammoth bones have also been found outside the recess, at the base of the cliff, which was a likely place of death for mammoths. But the scene of the crime was probably 50 metres above. This is not the only place on Earth in which such a scenario has been uncovered, and the pattern emerging has led to a theory of how mammoths were hunted.
It is unlikely that mammoths simply wandered too close to the edges of cliffs, rather that 50,000 years ago hyenas hunted on open plains, some of which ended abruptly at cliff edges. Pursued by hyenas towards the cliffs, it is probable that on occasion a mammoth tumbled over the edge. Mammoth bones at the foot of cliffs suggest that sometimes they fell over the edge, and the piles of bones from many individuals would seem to be more than a coincidence. So the hyenas living in the den below the cliff would be ideally placed to consume the carcasses. Theoretically, this is a good hunting strategy, and one that can be deduced from the fossil record and geological formation. But again, the real evidence that ancient hyenas ate mammoths lies in the teeth marks in the bones.
Back to the CambrianDiscussion so far has covered events that took place long after the Cambrian period, but what about the Cambrian itself? Are the equivalent of teeth and teeth marks recorded in the Cambrian fossils? We can turn to the Burgess Shale for evidence for the last time.
In the Burgess Shale are found groups of animals that exist today as predators. The jellyfish-like comb jelly
Fasciculus
would have pulsated through the shallow Cambrian seas swallowing any suitable prey in its path. The priapulid worms
Ancalagon
,
Louisella
,
Ottoia
and
Selkirkia
would have lain buried in the Cambrian sea floor waiting for some
unsuspecting creature to pass over their tubes. To tread near the entrance of these shafts would have been like stepping on a land mine to most Cambrian animals. The mouths of comb jellies are simply apertures, whereas the mouthparts of priapulids consist of a reversible proboscis, or mouth, and âlips'. This is obviously more complex and leaves its mark in the fossil record. The proboscis can be withheld inside the head, then extended out into the environment by a process of turning inside out. In this extended position, the lips are revealed at the extremity, along with rows of spines and teeth capable of ensnaring prey. When hooking is complete, the whole proboscis would be inverted back into the head, taking the ambushed prey with it. Most of the Burgess bristle worms also possessed a reversible proboscis, although not one so heavily laden with offensive spines. That is because most Burgess bristle worms fed on organic particles in the sediment or were scavengers on carcasses. But a more complex array of feeding parts leaves greater signs of predation in the fossil record, and such an array belonged to the active predators - those that actively hunted their prey.
Fasciculus
would have pulsated through the shallow Cambrian seas swallowing any suitable prey in its path. The priapulid worms
Ancalagon
,
Louisella
,
Ottoia
and
Selkirkia
would have lain buried in the Cambrian sea floor waiting for some
unsuspecting creature to pass over their tubes. To tread near the entrance of these shafts would have been like stepping on a land mine to most Cambrian animals. The mouths of comb jellies are simply apertures, whereas the mouthparts of priapulids consist of a reversible proboscis, or mouth, and âlips'. This is obviously more complex and leaves its mark in the fossil record. The proboscis can be withheld inside the head, then extended out into the environment by a process of turning inside out. In this extended position, the lips are revealed at the extremity, along with rows of spines and teeth capable of ensnaring prey. When hooking is complete, the whole proboscis would be inverted back into the head, taking the ambushed prey with it. Most of the Burgess bristle worms also possessed a reversible proboscis, although not one so heavily laden with offensive spines. That is because most Burgess bristle worms fed on organic particles in the sediment or were scavengers on carcasses. But a more complex array of feeding parts leaves greater signs of predation in the fossil record, and such an array belonged to the active predators - those that actively hunted their prey.
Other books
King of the Mountain by Fran Baker
19 Headed for Trouble by Suzanne Brockmann
Warning by Sophie Cunningham
Playing Dirty: Windy City Kink, Book 3 by Kelly Jamieson
Finding Her A-Muse-Ment by Rebecca Royce
PARANORMAL ROMANCE: The New Couple in Town: Don’t Let Them In by Natalia Napoli
The Riding Master by Alexandrea Weis
Is It Just Me? by Miranda Hart
A Stranger in Mayfair by Charles Finch
Sytravious: The Lost Warlock Of Moruz (The Oathbreaker Book 1) by Acevedo, E.B.