In The Blink Of An Eye (40 page)
Read In The Blink Of An Eye Online
Authors: Andrew Parker
Figure 8.5
Photograph of a trilobite when rolled up - âhead' spines can be seen projecting from the body. When the trilobite is flat, as we usually view trilobites, these spines lie flush with the body.
Photograph of a trilobite when rolled up - âhead' spines can be seen projecting from the body. When the trilobite is flat, as we usually view trilobites, these spines lie flush with the body.
The sponges
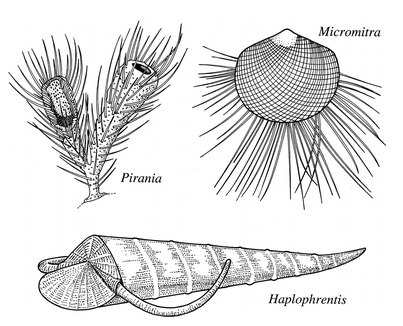
Choia
,
Halichondrites
,
Pirania
and
Wapkia
of the Burgess Shale contained spicules that not only provided a supporting lattice, but also projected into the environment as deadly blades. Burgess priapulid worms had spines in the region of their mouths for feeding, but also on other parts of their bodies, where they took their most fearsome forms. Like most lamp shells, the Burgess hyolith
Haplophrentis
completely closed shop by surrounding its entire body with exceptionally hard armour. The Burgess echinoderms, relatives of starfish today, similarly revealed no soft parts to a passing predator.
And finally we can consider
Micromitra
, where a shell as hard as a mussel's was obviously not enough to escape predation - it further evolved long spines around its edge.
Choia
,
Halichondrites
,
Pirania
and
Wapkia
of the Burgess Shale contained spicules that not only provided a supporting lattice, but also projected into the environment as deadly blades. Burgess priapulid worms had spines in the region of their mouths for feeding, but also on other parts of their bodies, where they took their most fearsome forms. Like most lamp shells, the Burgess hyolith
Haplophrentis
completely closed shop by surrounding its entire body with exceptionally hard armour. The Burgess echinoderms, relatives of starfish today, similarly revealed no soft parts to a passing predator.
And finally we can consider
Micromitra
, where a shell as hard as a mussel's was obviously not enough to escape predation - it further evolved long spines around its edge.
An earlier lamp shell,
Mickwitzia
, may have taken protection a stage further.
Mickwitzia
possibly employed chemical defences - it squirted toxins through holes in its shell. The evidence for this derives from the other shelly fossils found with
Mickwitzia
, which all exhibited boreholes made by predators.
Mickwitzia
, on the other hand, was always borehole free. To conclude, all of this evidence can mean only one thing - that animals possessed protection against predators in the Cambrian.
Mickwitzia
, may have taken protection a stage further.
Mickwitzia
possibly employed chemical defences - it squirted toxins through holes in its shell. The evidence for this derives from the other shelly fossils found with
Mickwitzia
, which all exhibited boreholes made by predators.
Mickwitzia
, on the other hand, was always borehole free. To conclude, all of this evidence can mean only one thing - that animals possessed protection against predators in the Cambrian.
The hard parts described so far all evolved at one point in time. This evolution was the Cambrian explosion - all animal phyla suddenly evolved their hard parts simultaneously between 543 and 538 million years ago. As mentioned already, hard parts can have functions other than to provide protection against predators, but it would appear extremely coincidental for all phyla to evolve hard parts at precisely the same time to provide strength or as a barrier against osmotic stress. Multicelled animals from different phyla had been around, in soft-bodied form, for 100 million years or so beforehand. And as established in Chapter 1, physical environmental conditions that could have demanded hard parts were not the cause of the Cambrian explosion. Now it becomes important to chart the original appearance of predators, particularly the highly active forms. This will be investigated as soon as all the clues from the Cambrian have been gathered.
The fact that all Burgess arthropods possessed protective spines, or some form of protection against attack, means they were not only predators, they were also prey. With the exception of the top predator
Anomalocaris
(which lacks protective spines) it is not surprising we could not deduce whether most Burgess animals with eyes were predators or prey based on their optics. In fact the ambiguity in the optical data supports the idea that most Cambrian animals in the open water were looking out for both prey and predators. With
Anomalocaris
a common menace in the Cambrian, the first rule was to stay alive, which meant keeping a lookout for the big-eyed giant. Cambrian eyes must have been adapted for scanning the complete environment, and any modification to this must have been slight due to the wrath of
Anomalocaris
and other highly mobile predators. That, as it happens, is exactly what we found in the Burgess eyes - adaptation for 360° of vision, with minor directional qualities.
Anomalocaris
(which lacks protective spines) it is not surprising we could not deduce whether most Burgess animals with eyes were predators or prey based on their optics. In fact the ambiguity in the optical data supports the idea that most Cambrian animals in the open water were looking out for both prey and predators. With
Anomalocaris
a common menace in the Cambrian, the first rule was to stay alive, which meant keeping a lookout for the big-eyed giant. Cambrian eyes must have been adapted for scanning the complete environment, and any modification to this must have been slight due to the wrath of
Anomalocaris
and other highly mobile predators. That, as it happens, is exactly what we found in the Burgess eyes - adaptation for 360° of vision, with minor directional qualities.
As for who ate who exactly, we can assume that the larger swimming forms preyed upon both the smaller swimming forms and the soft-bodied bottom-dwellers. But in addition to the telltale scars of
Anomalocaris
, there are other signs of predation in action where we can solve this problem more precisely. Fragments of the Burgess hyolith
Haplophrentis
have been discovered in the guts of thirty individuals of
Ottoia
, a priapulid worm, and also in the gut of the large arthropod
Sidneyia
. In that same gut of
Sidneyia
have been found seed-shrimps and trilobites -
Sidneyia
could feed on hard-shelled animals. And a closer inspection of the gut of one
Ottoia
revealed part of another
Ottoia
- so this priapulid worm was a cannibal.
Anomalocaris
, there are other signs of predation in action where we can solve this problem more precisely. Fragments of the Burgess hyolith
Haplophrentis
have been discovered in the guts of thirty individuals of
Ottoia
, a priapulid worm, and also in the gut of the large arthropod
Sidneyia
. In that same gut of
Sidneyia
have been found seed-shrimps and trilobites -
Sidneyia
could feed on hard-shelled animals. And a closer inspection of the gut of one
Ottoia
revealed part of another
Ottoia
- so this priapulid worm was a cannibal.
One fossil I picked up from the display table at the Burgess quarry was also interesting from this respect. This was the shrimp-like crustacean
Canadaspis
. . . and the tiny trilobite
Ptychagnostus
. The trilobite lay within the rounded head shield of
Canadaspis
and was probably its dinner. Other small trilobites have been found within the head shields of other Burgess arthropods, and it is possible that they were parasites. Since
Ptychagnostus
is found commonly in isolation, and probably lived in mid-water, maybe it was both parasite and prey.
Canadaspis
. . . and the tiny trilobite
Ptychagnostus
. The trilobite lay within the rounded head shield of
Canadaspis
and was probably its dinner. Other small trilobites have been found within the head shields of other Burgess arthropods, and it is possible that they were parasites. Since
Ptychagnostus
is found commonly in isolation, and probably lived in mid-water, maybe it was both parasite and prey.
It is worth a closer look at this situation from another perspective.
Canadaspis
has eyes whereas
Ptychagnostus
does not.
Canadaspis
and
Ptychagnostus
evolved to gamble on different aspects of The Laws of Life.
Canadaspis
placed its chips on âeating',
Ptychagnostus
on breeding.
Ptychagnostus
was extremely common in the Cambrian, whereas
Canadaspis
, and all other large Cambrian predators, was far less numerous.
Ptychagnostus
as a species was obviously prepared for predation - its survival was dependent on numbers. In other words,
Ptychagnostus
must have evolved a successful breeding strategy, so there were more individuals living in the water than could be consumed by predators - the strategy taken by krill in response to baleen whales. But this bottomless-pit-of-food scenario was not so accommodating to
Canadaspis
and its fellow predators. There remained the simple matter of âsearch and destroy'.
Canadaspis
has eyes whereas
Ptychagnostus
does not.
Canadaspis
and
Ptychagnostus
evolved to gamble on different aspects of The Laws of Life.
Canadaspis
placed its chips on âeating',
Ptychagnostus
on breeding.
Ptychagnostus
was extremely common in the Cambrian, whereas
Canadaspis
, and all other large Cambrian predators, was far less numerous.
Ptychagnostus
as a species was obviously prepared for predation - its survival was dependent on numbers. In other words,
Ptychagnostus
must have evolved a successful breeding strategy, so there were more individuals living in the water than could be consumed by predators - the strategy taken by krill in response to baleen whales. But this bottomless-pit-of-food scenario was not so accommodating to
Canadaspis
and its fellow predators. There remained the simple matter of âsearch and destroy'.
The open water is three-dimensional. One is less likely to bump into an animal in the open water than on the sea floor, which is a two-dimensional environment. The ocean is vast, and
Ptychagnostus
mobile. To catch
Ptychagnostus
one must have the ability to find it and swim after it. Thanks to the Cambrian explosion, the strong, skeletonised limbs with internal muscles achieved the mobility required. And like dragonflies in the air today, the eyes of
Canadaspis
gave it the means to find. Here we are building a picture of how life functioned in the Cambrian, and the rules are similar to those today. In Chapter 3 it was revealed that eyes and armour are not directly related in that
species with eyes do not always have armour, and vice versa. However, there may be a link in behaviour . . . and evolution.
Ptychagnostus
mobile. To catch
Ptychagnostus
one must have the ability to find it and swim after it. Thanks to the Cambrian explosion, the strong, skeletonised limbs with internal muscles achieved the mobility required. And like dragonflies in the air today, the eyes of
Canadaspis
gave it the means to find. Here we are building a picture of how life functioned in the Cambrian, and the rules are similar to those today. In Chapter 3 it was revealed that eyes and armour are not directly related in that
species with eyes do not always have armour, and vice versa. However, there may be a link in behaviour . . . and evolution.
Chapter 1 introduced an old idea for the cause of the Cambrian explosion, one reworked recently by Mark McMenamin and Dianna Schulte McMenamin - that a food web was developed at the beginning of the Cambrian, where every species had its own predators and food. But can entire food webs simply spring up out of nowhere? Or is there a factor that triggers a chain reaction, ending in the formation of a full-blown food web? The range of animals living just after the Cambrian explosion, with their diversity of shapes and sizes, suggests a mature food web was in place at that time. But did this food web mature quickly, or instantaneously as the Cambrian enigma demands? Or did it assemble gradually, beginning in the Precambrian? These questions indicate that our Cambrian jigsaw puzzle is nearly complete.
The McMenamins resurrected a century-old idea that animals developed shells as shields against predators, a fact we too have established. In this chapter, predators are emerging as hugely important factors in the way life works today and how it did in the past. But when did the remaining feeding modes within food webs first appear? This question may be irrelevant to our overall quest if food webs, and consequently The Laws of Life, were established
before
the Cambrian. It would seem appropriate for this chapter to end in the manner of the previous one, in a search for the
beginning
of predators on Earth.
In the original line of firebefore
the Cambrian. It would seem appropriate for this chapter to end in the manner of the previous one, in a search for the
beginning
of predators on Earth.
Journeying beyond the Cambrian explosion and into the ârelative unknown' that is the Precambrian, the first port of call is the age of Ediacara. The Ediacaran suite of life forms is best represented by the original finds from South Australia, around 565 million years old, although the same organisms existed right up until the Cambrian explosion itself, when they disappeared without trace. But while they existed, they did exhibit a variety of lifestyles. We know this from the shapes of the life forms themselves and from their trace fossils - footprints and their equivalents.
The Cambrian was once described as a peaceful time, but we have now established this is not true. In fact we know now that predators existed even before. In the Precambrian there were jellyfish pulsating through mid-water, and relatives of the Portuguese man-of-war floating on the surface. Any creature that accidentally encountered the stinging tentacles of these animals would have instantly become their prey. On the sea floor were anemone-like creatures with their stinging tentacles waving expectedly upwards. And then there was Precambrian prey. In some cases the Ediacaran predators would have preyed upon each other. But there were also flat, worm-shaped animals that probably undulated their bodies to propel them through the water - and sometimes into the lions' den. Occasionally they would have propelled themselves into the nets of stinging tentacles, cast hopefully into the water.
Although the word âhopefully' infers personality in these primitive forms, it
is
appropriate in that Precambrian predation was a comparatively random process. There was no
Anomalocaris
with its advanced detection system and search-and-destroy capabilities. All that patrolled the Precambrian water were the stinging nets of jellies. But in the jelly's favour, the prey could not sense them coming either.
is
appropriate in that Precambrian predation was a comparatively random process. There was no
Anomalocaris
with its advanced detection system and search-and-destroy capabilities. All that patrolled the Precambrian water were the stinging nets of jellies. But in the jelly's favour, the prey could not sense them coming either.
This last statement may not be strictly true. Although too small to be recorded as fossils, Ediacaran organisms surely possessed sense organs of some kind. They may have sensed vibrations in the water, based on movements of tiny hairs on their skin, which could signal the advance of a barrage of stinging cells. Indeed, the probable relatives of some Ediacarans today are endowed with hairs of this type. But detection in the Precambrian would have been possible only in close encounters. And selection for a more advanced sensing system would have been minimised by the generally slow speed of the advancing predators. This was a kitten-and-mouse game, in comparison with the cat-and-mouse Cambrian.
On the sea floor the threat of predation was no more severe . . . but did exist all the same. A worm-like animal called
Claudina
lived in the sediment just prior to the Cambrian, about 550 million years ago. It is known from precisely 524 fossils from Shaanxi province in China. The fossils are not of the animal itself, but of its tube - this is the first
animal known to possess hard parts. It appeared to have jumped the gun before the start of the Cambrian, at the same time demonstrating that environmental conditions were not completely restrictive for making hard parts before the great explosion.
Claudina
lived in the sediment just prior to the Cambrian, about 550 million years ago. It is known from precisely 524 fossils from Shaanxi province in China. The fossils are not of the animal itself, but of its tube - this is the first
animal known to possess hard parts. It appeared to have jumped the gun before the start of the Cambrian, at the same time demonstrating that environmental conditions were not completely restrictive for making hard parts before the great explosion.
Fourteen tubes of
Claudina
revealed boreholes - holes made by a predator on the sea floor in a successful attempt to consume the soft animal within. Stefan Bengtson of Uppsala University and Yue Zhao of the Chinese Academy of Geological Sciences, who found these fossils, believe the predator to have been a mollusc, possibly a relative of snails today. But in the Precambrian, molluscs, like most other animal phyla, looked like âworms', or rather had completely soft bodies. There was not even a hint that one day its descendants would carry around huge shells.
Claudina
revealed boreholes - holes made by a predator on the sea floor in a successful attempt to consume the soft animal within. Stefan Bengtson of Uppsala University and Yue Zhao of the Chinese Academy of Geological Sciences, who found these fossils, believe the predator to have been a mollusc, possibly a relative of snails today. But in the Precambrian, molluscs, like most other animal phyla, looked like âworms', or rather had completely soft bodies. There was not even a hint that one day its descendants would carry around huge shells.
The holes in the tubes of
Claudina
provide the first definitive evidence of predation on Earth. And it seems that what can be best described as âinactive predation' was common in the Precambrian. Although, based on the lack of armour worn in the Precambrian, this type of predation obviously did not present a strong selection pressure for counter-predatory measures. It did not provide the stimulus for hard, protective parts.
Claudina
provide the first definitive evidence of predation on Earth. And it seems that what can be best described as âinactive predation' was common in the Precambrian. Although, based on the lack of armour worn in the Precambrian, this type of predation obviously did not present a strong selection pressure for counter-predatory measures. It did not provide the stimulus for hard, protective parts.
In particular, there was one interesting soft-bodied animal that roamed the Precambrian sea floor. In 1984, petroleum companies were exploring parts of southern Morocco and eastern Siberia. They drilled vertically into the ground and removed cores - long, thin cylinders of rock that revealed the layers of sediment built up over 600 million years, while these areas were underwater. As expected, rocks that formed just before the Cambrian showed signs of stromatolites. But there were further, unusual layers just above the stromatolites. At the time they were termed âthrombolites' and were assumed to be the result of grazing by soft-bodied arthropods, including âproto-trilobites'. Indeed, the first signs of trilobites, the first hard parts of any type, were found some tens of metres above the lowest thrombolites. Along with trace fossils of soft-bodied arthropods, this is an important clue in piecing together a picture of the ancestors of arthropods. But most enticing is the term âproto-trilobite'. Did trilobites as such exist without their armour before the Cambrian explosion? In 1991 this
question was answered. Incredibly, a new expedition to the original Ediacaran site, the Ediacaran Hills in South Australia, yielded a soft-bodied trilobite.
question was answered. Incredibly, a new expedition to the original Ediacaran site, the Ediacaran Hills in South Australia, yielded a soft-bodied trilobite.
Other books
Burying the Honeysuckle Girls by Emily Carpenter
Vanishing Point by Alan Moore
Five Go Down to the Sea by Enid Blyton
HIGH TIDE by Miller, Maureen A.
Chosen by P.C. Cast and Kristin Cast, Kristin Cast
Dirty Little Thing by Sara Brookes
How to Survive Middle School by Donna Gephart
Second Chances by Charity Norman
Prince of Fire and Ashes: Book 3 of the Tielmaran Chronicles by Reimann, Katya
Outcast (SEAL Team: Disavowed Book 2) by Laura Marie Altom