In The Blink Of An Eye (39 page)
Read In The Blink Of An Eye Online
Authors: Andrew Parker
A Cambrian arrow worm has been uncovered from the Stephen Formation of the Burgess site. This was buried at a deeper water location than most of the other Burgess fossils, but it was an active swimmer so may also have inhabited shallow-water sites. Interestingly, just as arrow worms are today, then it was also a predator. We know this because it possessed the characteristic arrow worm mouth spines, the tools that grasp prey in mid-water. The prey in this case would have been small and planktonic, but other active predators in Cambrian seas were large, and their grasping tools and mouthparts were fearsome.
The most memorable fossil I examined at the Burgess quarry, protected under its plastic sheet, was a specimen of
Anomalocaris
. One look at its grasping forelimbs and the word âpredator' springs immediately to mind.
Anomalocaris
was widespread at least between 525 and 515 million years ago, when it was the number one predator. At up to 2 metres long it was certainly the largest animal of its time.
Anomalocaris
. One look at its grasping forelimbs and the word âpredator' springs immediately to mind.
Anomalocaris
was widespread at least between 525 and 515 million years ago, when it was the number one predator. At up to 2 metres long it was certainly the largest animal of its time.
Recently, the Japanese Broadcasting Corporation NHK made a full-size model of
Anomalocaris
for a documentary series. The overlapping
flaps along the side of the animal were undulated in a wave-like manner, and the model emerged highly manoeuvrable, like cuttlefishes today. It could move forwards, backwards or simply hover in mid-water. So the Burgess and a similar Chinese
Anomalocaris
species were able to actively swim after their prey. An Australian species, on the other hand, was more cumbersome and probably combed the mud for its prey. But all species of
Anomalocaris
benefited from the same type of circular mouth, a collection of hard plates that open and close like the iris of a camera, with a circular array of teeth inside. The aperture itself was rectangular and could not be closed - the teeth did not meet in the middle. Rather the mouth was opened further to admit prey, then the hard plates were pulled together to draw the prey into the mouth. This action would have cracked or even broken the armour of arthropods. Unfortunately, before this reconstruction of a large, fearsome arthropod,
Anomalocaris
and its various parts had to pass through other interpretations. Throughout the history of palaeontology it has been a jellyfish, a sea cucumber, a bristle worm, a sponge and a shrimp. Sometimes it is worth keeping digging.
Anomalocaris
for a documentary series. The overlapping
flaps along the side of the animal were undulated in a wave-like manner, and the model emerged highly manoeuvrable, like cuttlefishes today. It could move forwards, backwards or simply hover in mid-water. So the Burgess and a similar Chinese
Anomalocaris
species were able to actively swim after their prey. An Australian species, on the other hand, was more cumbersome and probably combed the mud for its prey. But all species of
Anomalocaris
benefited from the same type of circular mouth, a collection of hard plates that open and close like the iris of a camera, with a circular array of teeth inside. The aperture itself was rectangular and could not be closed - the teeth did not meet in the middle. Rather the mouth was opened further to admit prey, then the hard plates were pulled together to draw the prey into the mouth. This action would have cracked or even broken the armour of arthropods. Unfortunately, before this reconstruction of a large, fearsome arthropod,
Anomalocaris
and its various parts had to pass through other interpretations. Throughout the history of palaeontology it has been a jellyfish, a sea cucumber, a bristle worm, a sponge and a shrimp. Sometimes it is worth keeping digging.
The five-eyed
Opabinia
is another obvious predator with its moveable, snapping mouthpart.
Opabinia
was just as manoeuvrable in the water as
Anomalocaris
- actually they were probably related. The snapping mouthpart of
Opabinia
probably represents the grasping forelimbs of
Anomalocaris
- twisted 90° with their bases elongated into a tube. Based on the shapes of the bodies and limbs alone, it would appear that the list of active predatory forms represented in the Burgess Shale is long.
Opabinia
is another obvious predator with its moveable, snapping mouthpart.
Opabinia
was just as manoeuvrable in the water as
Anomalocaris
- actually they were probably related. The snapping mouthpart of
Opabinia
probably represents the grasping forelimbs of
Anomalocaris
- twisted 90° with their bases elongated into a tube. Based on the shapes of the bodies and limbs alone, it would appear that the list of active predatory forms represented in the Burgess Shale is long.
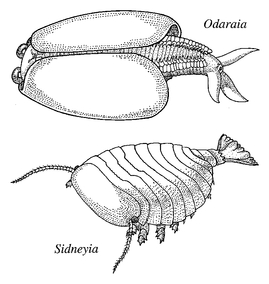
Most of the large arthropods of the Burgess Shale were certainly predators, actively hunting their prey in mid-water. Some, like
Odaraia
, do not have large grasping limbs and would have preyed upon shoals of small floating or swimming organisms. Others, like
Sanctacaris
and
Sidneyia
, were armed with a barrage of spines and claws, and would have been formidable predators to most Burgess organisms. But what about the best represented group of arthropods in the Cambrian - the trilobites?
Odaraia
, do not have large grasping limbs and would have preyed upon shoals of small floating or swimming organisms. Others, like
Sanctacaris
and
Sidneyia
, were armed with a barrage of spines and claws, and would have been formidable predators to most Burgess organisms. But what about the best represented group of arthropods in the Cambrian - the trilobites?
Some Cambrian trilobites had sizeable digestive chambers for the initial processing of food. These, surely, were predators - they needed to
store single, large food items at a particular time. Such large digestive chambers are not found in detritus-feeding trilobites, those species that combed the sea floor for particles of organic matter. There were indeed trilobites which employed such feeding methods, and others that were planktivores, filter feeders . . . some even cultured bacteria to provide a meal. Much of the evidence for this comes from the specific shapes of the fossils themselves. For instance, trilobite expert Richard Fortey of the Natural History Museum in London noticed the bulging sides and reduced mouthparts of one trilobite. From this he understood that food was absorbed through the gills along the sides of the body, derived from bacterial colonies living there. Today, crustaceans living at mid-oceanic ridges and hydrothermal vents obtain nutrients from similar bacteria housed in their gill structures. As further supporting evidence, Fortey's trilobite inhabited a similar environment.
store single, large food items at a particular time. Such large digestive chambers are not found in detritus-feeding trilobites, those species that combed the sea floor for particles of organic matter. There were indeed trilobites which employed such feeding methods, and others that were planktivores, filter feeders . . . some even cultured bacteria to provide a meal. Much of the evidence for this comes from the specific shapes of the fossils themselves. For instance, trilobite expert Richard Fortey of the Natural History Museum in London noticed the bulging sides and reduced mouthparts of one trilobite. From this he understood that food was absorbed through the gills along the sides of the body, derived from bacterial colonies living there. Today, crustaceans living at mid-oceanic ridges and hydrothermal vents obtain nutrients from similar bacteria housed in their gill structures. As further supporting evidence, Fortey's trilobite inhabited a similar environment.
Using crustaceans as their modern-day representatives, it would seem
that most trilobites were predator-scavengers. That is, they fed on the bodies of other multicelled animals, either living or dead. Their heavily spined, robust limbs could have had no other purpose but to grasp and tear apart whole animals. As will be examined in more detail at the end of this chapter, the majority of
early
trilobites were active predators - they moved rapidly to hunt their prey. Further evidence to support this view is sealed within fossils of the Naraoids.
that most trilobites were predator-scavengers. That is, they fed on the bodies of other multicelled animals, either living or dead. Their heavily spined, robust limbs could have had no other purpose but to grasp and tear apart whole animals. As will be examined in more detail at the end of this chapter, the majority of
early
trilobites were active predators - they moved rapidly to hunt their prey. Further evidence to support this view is sealed within fossils of the Naraoids.
Naraoids were a sister group of trilobites - they were their closest relatives and bore a physical resemblance. Naraoids too possessed very spiny and formidable-looking limbs, in addition to fang-bearing mouthparts. They probably fed on worms and other soft-bodied creatures. But Naraoids were different from trilobites in one respect - they had relatively soft bodies. The upper surface of their exoskeleton was not calcified like that of a trilobite, but was only organically strengthened. For this reason, to support the massive spiny limbs the upper surface of the body could not have been jointed like that of a trilobite - it would have been too weak. The upper body surface is a point for muscle attachments and is comparable to the supporting walls of a house. So the predatory limbs of Naraoids came at quite a cost to their bodies.
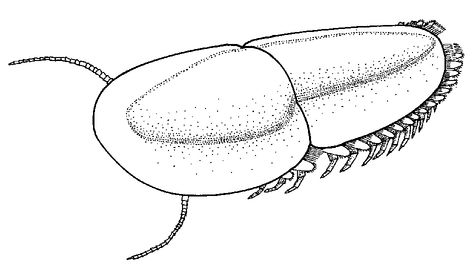
Naraoids evolved from the ancestor of the trilobites before the exoskeleton became strengthened with calcium carbonate, or calcite. Naraoids are often found distorted in their burial grounds as a result of their weak exoskeletons, and they also missed out on eyes. The important evolutionary history of Naraoids and trilobites will be considered further in this chapter, but first we should examine the remaining evidence for predation in the Cambrian. So far we have looked at eyes, feeding apparatus and digestive systems. Now we should look for teeth marks in the prey.
The display of Burgess Shale fossils at the information centre in the Canadian town of Field contains an interesting trilobite. Although most fossils here are remarkably complete and favourably orientated in the rock, and include some of the best examples of the Burgess species, a specimen of the trilobite
Olenoides
is particularly noteworthy. A large part of its body is missing, but the regularity of the semicircular omission suggests this was not an artefact of preservation. It could be only one thing - a bite mark. A large Cambrian predator bit this trilobite - it was Cambrian prey.
Olenoides
is particularly noteworthy. A large part of its body is missing, but the regularity of the semicircular omission suggests this was not an artefact of preservation. It could be only one thing - a bite mark. A large Cambrian predator bit this trilobite - it was Cambrian prey.
Many other Cambrian trilobites have been found with scars, signs of attack by a predator sustained while still alive. These wounds proved not to be fatal because of the animals' ability to heal. This is an interesting concept in itself. Cambrian trilobites were well prepared for attack not only in their protective armour but also in their ability quickly to seal the newly exposed body sections - they could form calluses. Human skin is thin and can be easily cut. For this reason, our blood has the ability to coagulate and seal up broken blood vessels, preventing blood loss and infection. Arthropod exoskeletons, on the other hand, are tough and designed to withstand the rigours of their hosts' lifestyles . . . except when they are heavily attacked. The self-healing of Cambrian trilobites indicates they were so prone to attack that predation had certainly been a selection pressure during their evolution. Today animals can be found with hard shells that have functions other than to protect them against predators, such as providing support for tissues. But not only had Cambrian trilobites evolved armour, they had also evolved a self-healing mechanism to function in the event of attack by predators. Their hard shells had a role in protection against predators from the beginning.
There have been so many Cambrian trilobites found with bite marks that a theory of âhandedness' has been suggested. In a large sample of trilobites, seventy-seven specimens had sustained injuries of unknown origin, perhaps caused by accidents during moulting or mating, but eighty-one specimens revealed injuries caused by predatory attacks. Researchers at Ohio State University found that 70 per cent of all scars left by predators were on the right side of the trilobites. It is thought that trilobites, their predators, or more likely both, tended to favour one side. Trilobites probably veered to one side in an attempt to evade an attacker. Also, predators probably tended to attack from the same side. Such asymmetrical behaviour is commonly seen today - a horse tends to turn its head to the left and 90 per cent of humans are right handed. But of most relevance to this chapter is the shape of the trilobite scars, whether on the left or the right side of the body. Many were W-shaped, conspicuously matching the size and shape of the iris-forming, triangular mouth plates of
Anomalocaris
.
Anomalocaris
.
With the exception of the trilobite-like
Naraoia
, all the Burgess arthropods were protected within armour. They possessed head shields that were sometimes further protected by solid bumps or spines. Many trilobites possessed large spines - the defensive role of these becomes obvious when trilobites are considered in their curled-up posture. Here the animal is transformed into a hard ball with projecting spines. Furthermore, the spines are sometimes quite elaborate, with serrations and spikes.
Naraoia
, all the Burgess arthropods were protected within armour. They possessed head shields that were sometimes further protected by solid bumps or spines. Many trilobites possessed large spines - the defensive role of these becomes obvious when trilobites are considered in their curled-up posture. Here the animal is transformed into a hard ball with projecting spines. Furthermore, the spines are sometimes quite elaborate, with serrations and spikes.
Long, sharp spines can be found on many hard exoskeletons, but also protecting softer, more fragile bodies like that of the Burgess lace âcrab',
Marrella
. In fact the use of armoured spines to protect soft bodies was employed by animals from a range of diverse phyla. There was the velvet worm
Hallucigenia
with its soft body protected by long, upwardly projecting spines. The bristle worms are more classical cases of this phenomenon;
Canadia
, for instance, wore a coat of spines projecting upwards and sideways. It is thought that bristle worms and their relatives in freshwater today independently evolved spines for defence purposes. Such convergence suggests that this means of protection is a good one. The epitome of protection within the Burgess bristle worms, however, was to be found in
Wiwaxia
, an oval-shaped
animal not only completely covered by overlapping shields but also with long swords projecting outwards that made even
Hallucigenia
look like easy pickings. Halkieriids are possible ancestors of, and bore a close resemblance to,
Wiwaxia
- they possessed a similar means of protection in their chain-mail coat of shields.
Marrella
. In fact the use of armoured spines to protect soft bodies was employed by animals from a range of diverse phyla. There was the velvet worm
Hallucigenia
with its soft body protected by long, upwardly projecting spines. The bristle worms are more classical cases of this phenomenon;
Canadia
, for instance, wore a coat of spines projecting upwards and sideways. It is thought that bristle worms and their relatives in freshwater today independently evolved spines for defence purposes. Such convergence suggests that this means of protection is a good one. The epitome of protection within the Burgess bristle worms, however, was to be found in
Wiwaxia
, an oval-shaped
animal not only completely covered by overlapping shields but also with long swords projecting outwards that made even
Hallucigenia
look like easy pickings. Halkieriids are possible ancestors of, and bore a close resemblance to,
Wiwaxia
- they possessed a similar means of protection in their chain-mail coat of shields.

Other books
The Orc's Tale by Jonathan Moeller
Genesis - the Battle Within (Pillars of Creation Book 1) by Tucker, David
For the Love of Old Bones - and other stories (Templar Series) by Jecks, Michael
Against the Wall (Stoddard Art School Series Book 3) by Lisa A. Olech
Pulse (Contemporary new adult/college romance) (Club Grit Trilogy) by Jaxsen, Brooke
BlackmailedbyHisRival by Adriana Rossi
The Dead Media Notebook by Bruce Sterling, Richard Kadrey, Tom Jennings, Tom Whitwell
Born Under Punches by Martyn Waites
Somerset by Leila Meacham
Los exploradores de Pórtico by Frederik Pohl