What Technology Wants (18 page)
Read What Technology Wants Online
Authors: Kevin Kelly
The constraints of physics, chemistry, and geometry have governed life from its origins onwardâand even into the technium. “Underlying all the diversity of life is a finite set of natural forms that will recur over and over again anywhere in the cosmos where there is carbon-based life,” claim biochemists Michael Denton and Craig Marshall. Evolution simply cannot make all possible proteins, all possible light-gathering molecules, all possible appendages, all possible means of locomotion, all possible shapes. Life, rather than being boundless and unlimited in every direction, is bounded and limited in many directions by the nature of matter itself.
I will argue that the same constraints bind technology. Technology is based on the same physics and chemistry as life, and more important, as the seventh, accelerated kingdom of life, the technium is bound by many of the same constraints guiding life's evolution. The technium can't make all imaginable inventions or all possible ideas. Rather, the technium is limited in many directions by the constraints of matter and energy. But the negative constraints of evolution are only half of its story.
The second great force pushing evolution on its immense journey is positive constraints that channel evolutionary innovation in certain directions. In tandem with the constraints of physical laws outlined above, the exotropy of self-organization steers evolution along a trajectory. While these internal inertias are immensely important in biological evolution, they are even more consequential in technological evolution. In fact, in the technium, self-generated positive constraints are more than half the story; they are the main event.
However, the existence of internal constraints guiding biological evolution is far from orthodoxy in biology today. The notion of directional evolution has a colorful history tainted by its association with a belief in a supernatural essence of life. While it is no longer associated with the supernatural today, the idea of directional evolution is now associated with the idea of “inevitable”âa concept that many modern scientists find intolerable in any form.
I would like to present the best case for a direction within biological evolution that our evidence so far will permit. It is a complicated story, vital not only for understanding biology but also for discerning the future of technology. Because if I can demonstrate that there is an internally generated direction within natural evolution, then my argument that the technium extends this direction is easier to see. So while I delve deeply into the forces driving life's evolution, this long explanation is really a parallel argument for the same kind of evolution within technology.
I begin this second half of the story with a reminder that this newly appreciated exotropic drive of evolution is not its only engine. Evolution has multiple drivers, including the physical constraints I earlier described. But in the current orthodox scientific understanding of evolution, change is attributed chiefly to one source: random variation. In the wilds of nature, reproducing survivors are naturally selected from inheritable random variations; therefore, in evolution there can only be random advance without direction. The key insight gained by the last three decades of research on complex adaptive systems offers a contrary view: that the
variation presented to natural selection is not always random
. Experiments show that “random” mutations are often not unbiased; instead, variation is governed by geometry and physics; and most important, variations are often shaped by the possibilities inherent in the recurring patterns of self-organization (a la whirlpool vortex).
variation presented to natural selection is not always random
. Experiments show that “random” mutations are often not unbiased; instead, variation is governed by geometry and physics; and most important, variations are often shaped by the possibilities inherent in the recurring patterns of self-organization (a la whirlpool vortex).
Once upon a time the notion of nonrandom variation was heresy, but as more and more biologists ran computer models, the idea that variation is not random became a scientific consensus among certain theoreticians. Self-regulating networks of genes (found in all chromosomes) favor certain kinds of complexes. “Some potentially useful mutations are so probable that they can be viewed as being encoded implicitly in the genome,” says biologist L. H. Caporale. Metabolic pathways in cells can autocatalyze themselves into a network and drift into self-preferred loops. This flips the traditional view. In the old view, the internal (the source of mutation) created change, while the external (the environmental source of adaptation) selected or directed it; in the new view, the external (physical and chemical constraints) creates forms, while the internal (self-organization) selects or directs them. And when the internal directs, it redirects to recurring forms. As the early paleontologist W. B. Scott put it, the complexity of evolution creates “inherited channels for preferred change.”
In the textbook version, evolution is a mighty force propelled by a single near-mathematical mechanism: inheritable random mutations selected by adaptive survival, also known as natural selection. The emerging modified view recognizes additional forces. It proposes that the creative engine of evolution stands on
three
legs: the adaptive (the classic agent), plus the contingent and the inevitable. (These three forces reappear in the technium as well.) We can describe these as three vectors of evolution.
three
legs: the adaptive (the classic agent), plus the contingent and the inevitable. (These three forces reappear in the technium as well.) We can describe these as three vectors of evolution.
The adaptive vector is the orthodox force that textbook theory teaches. Just as Darwin surmised, those organisms that adapt best to their environment survive to breed offspring. So any new strategies for survival in a changing environment, no matter where they come from, are selected over time and lead to a very fine fit for that species. The adaptive force is fundamental at all levels of evolution.
The second vector in evolution's triad is luck, or contingency. A lot of what happens in evolution comes down to the lottery, not adaptation of the superior. Much of the fine detail of speciation is a result of happenstance, some improbable trigger that leads a species down a contingent path. The individual speckles on a monarch butterfly's wings are not strictly adaptive, just plain chance. These random beginnings can eventually lead to completely unexpected designs later on. And these subsequent designs may be less complex or less elegant than their predecessors. In other words, many of the forms we see in evolution today are due to random contingencies in the past and don't follow a progressive sequence. If we rewind the tape of life's history and push start again, it will play out differently. (I should mention for the benefit of young readers that “rewinding the tape,” like “dialing the phone,” “filming a movie,” or “cranking the engine,” is a skeuonym, an expression left over from a technology no longer used. In this case, “rewinding a tape” means to rerun a sequence from the same starting point.)
Stephen Jay Gould, who introduced the trope of “rewinding the tape of life” in his seminal book
Wonderful Life,
makes an elegant case for the ubiquity of contingency in evolution. He based his argument on the evidence of a set of cryptic fossils of pre-Cambrian life found in the Burgess Shale in Canada. A young grad student named Simon Conway Morris spent years tediously dissecting these minute fossils under a microscope. After a decade of intense study Morris announced that the Burgess Shale contained a treasure trove of previously unknown biota, far more diverse in forms than life now. But this great ancient diversity of archetypes was decimated by unlucky disasters 530 million years ago, leaving further evolution with only a relative few basic organism typesâthe comparatively less varied world we see now. Superior designs were randomly eliminated. Gould interpreted this chancy decimation of older, greater diversity as a powerful argument for the rule of contingency and an argument against the idea of directionality in evolution. In particular he believed the evidence of the Burgess Shale demonstrated that human minds were not inevitable, because nothing in evolution was inevitable. At the close of his book, Gould concludes, “Biology's most profound insight into human nature, status, and potential lies in the simple phrase, the embodiment of contingency:
Homo sapiens
is an entity, not a tendency.”
Wonderful Life,
makes an elegant case for the ubiquity of contingency in evolution. He based his argument on the evidence of a set of cryptic fossils of pre-Cambrian life found in the Burgess Shale in Canada. A young grad student named Simon Conway Morris spent years tediously dissecting these minute fossils under a microscope. After a decade of intense study Morris announced that the Burgess Shale contained a treasure trove of previously unknown biota, far more diverse in forms than life now. But this great ancient diversity of archetypes was decimated by unlucky disasters 530 million years ago, leaving further evolution with only a relative few basic organism typesâthe comparatively less varied world we see now. Superior designs were randomly eliminated. Gould interpreted this chancy decimation of older, greater diversity as a powerful argument for the rule of contingency and an argument against the idea of directionality in evolution. In particular he believed the evidence of the Burgess Shale demonstrated that human minds were not inevitable, because nothing in evolution was inevitable. At the close of his book, Gould concludes, “Biology's most profound insight into human nature, status, and potential lies in the simple phrase, the embodiment of contingency:
Homo sapiens
is an entity, not a tendency.”
This phrase “entity not tendency” is the orthodoxy in evolutionary theory today: that inherent contingency and supreme randomness in evolution preclude tendencies in any direction. However, later research disproved the notion that the Burgess Shale contained as great a diversity as first believed, deflating Gould's conclusions. Simon Conway Morris himself changed his mind about his earlier radical classifications. It turns out many of the Burgess Shale organisms were not weird new forms but weird old forms, and so contingency was far less prevalent in macroevolution, and progress more likely. Curiously, over the years since Gould's influential book, Morris has become the chief paleontologist championing the idea of convergence, directionality, and inevitabilities in evolution. In hindsight what the Burgess Shale proves is that contingency is a significant force in evolution, but not the only one.
The third leg of evolution's tripod is structural inevitability, the very force denied by the current dogma of biology. Whereas contingency can be thought of as a “historical” force, that is, a phenomenon where history matters, the structural component of evolution's engine can be thought of as “ahistorical” in that it produces change independent of history. Run it again, and you get the same story. This aspect of evolution pushes inevitabilities. For instance, the defensive venomous sting has been evolved at least twelve times: in the spider, the stingray, the stinging nettle, the centipede, the stonefish, the honeybee, the sea anemone, the male platypus, the jellyfish, the scorpion, the cone-shell mollusk, and the snake. Its reappearance is due not to a common history but to a common matrix of life, and that common structure arises not from the outside environment but from the internal momentum of self-organized complexity. This vector is the exotropic force, the emergent self-organization that arises in a system as complex as evolutionary life. As described in previous chapters, complex systems acquire their own inertia, creating recurring patterns that the system tends to fall into. This emergent self-order steers the system to its own selfish interests, and in this way it engenders a direction to the ongoing process. This vector pushes the messiness of evolution toward certain inevitabilities.
Charted, the tripod of evolution might look like this:
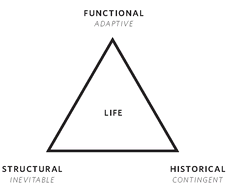
The Triad of Evolution.
The three evolutionary vectors in life. The bold name indicates the realm in which it operates and the italic name its consequence.
The three evolutionary vectors in life. The bold name indicates the realm in which it operates and the italic name its consequence.
All three dynamics are present in varying proportions at different levels in nature, counterbalancing and offsetting one another, combining to produce the history of each creature. A metaphor comes to mind that may help to untangle the three forces: The evolution of a species is like a meandering river as it carves away land. The detailed “particular-ness” of that river, the profile of its fine contours along the shore and bottom, comes from the vectors of adaptive mutations and contingency (never to be repeated), but the universal “riverness” form of the river (recurring in all rivers) as it is channeled in a valley comes from the internal gravity of convergence and emergent order.
For another example of contingent microdetails decorating inevitable macroarchetypes, consider the six separate dinosaur lineages that have followed the same morphological pathway in evolution. Over time each of the six dino lineages displayed a similar (inevitable) reduction in their side toes, an elongation of the long bones in their paws, and a shortening of their “fingers.” We might call this pattern part of “dinosaurness.” Because they are rehearsed in six lineages these archetypical structures are not merely random. Bob Bakker, the model for the dino guy in
Jurassic Park
and real-life dinosaur expert, claims, “This striking case of iterative parallelism and convergence [in the six dino lineages] . . . is a powerful argument that observed long-term changes in the fossil record are the result of directional natural selection, not a random walk through genetic drift.”
Jurassic Park
and real-life dinosaur expert, claims, “This striking case of iterative parallelism and convergence [in the six dino lineages] . . . is a powerful argument that observed long-term changes in the fossil record are the result of directional natural selection, not a random walk through genetic drift.”
Way back in 1897, paleontologist Henry Osborn, an early dinosaur and mammal expert, wrote: “My study of teeth in a great many phyla of Mammalia in past times has convinced me that there are fundamental predispositions to vary in certain directions; that the evolution of teeth is marked out beforehand by hereditary influences which extend back hundreds of thousands of years.”
It is important to outline what is “marked out beforehand.” In most cases, the details of life are contingent. The river of evolution determines only the broadest outlines of form. One might think of these as grand archetypes, for instance, tetrapods (four-leggedness), the snake form, eyeballs (spherical cameras), coiled guts, egg sacs, flapping wings, repeating segmented bodies, trees, puffballs, fingers. These are general silhouettes, not individuals. The biologist Brian Goodwin proposed that “all the main morphological features of organismsâhearts, brains, guts, limbs, eyes, leaves, flowers, roots, trunks, branches, to mention only the obvious onesâare the emergent results of morphogenic principles” and would reappear if the tape of life was rewound. Like other recurring archetypes, they are patterns your brain perceives without your even noticing it. “Oh, it's a clam,” your mind says to itself, letting you fill in the particulars of color, texture, and individual species. The “clam” formâtwo concave hemispheres hinged to closeâis the recurring archetype, the determined form.
Other books
Academ's Fury by Jim Butcher
The Rising Tide: A Novel of World War II by Jeff Shaara
Untamed (A Bad Boy Secret Baby Romance) by Emilia Kincade
Awake and Alive by Garrett Leigh
Only for You by Valentine, Marquita
The Love Letter by Matthews, Erica
Fear (June Hunt Hope for the Heart) by Hunt, June
i de1359f7e9a78273 by Unknown
Duke (Aces MC Series Book 2) by Foster, Aimee-Louise
Squirrel World by Johanna Hurwitz